- Физиология нормального сна

Содержание
- 2. «Кто познает тайну сна – познает тайну мозга». — МИШЕЛЬ ЖУВЕ (1925-2017)
- 3. Сон – это особое генетически детерминированное состояние организма гомеотермных (теплокровных) животных, характеризующееся закономерной последовательной сменой определенных
- 4. Цикл «бодрствование – сон» В регуляции цикла «бодрствование-сон» на системном уровне принимают участие 4 группы механизмов
- 5. Механизмы поддержания бодрствования (1) На уровне ромбовидного мозга: нейроны синего пятна (норадреналин) нейроны педункулопонтинной и латеродорзальной
- 6. Под воздействием восходящей активации нейроны коры переходят в состояние так называемой «тонической деполяризации», сдвигая свой мембранный
- 7. Механизмы медленного сна Основное скопление тормозных нейронов отмечено в вентролатеральной и срединной частях преоптической области переднего
- 8. Роль аденозина Одна из гипотез связывает причину медленного сна с постепенным накоплением в ходе длительного бодрствования
- 9. Стадии медленного сна 1 стадия (дремота): 5-10 мин снижается мышечная активность глаза могут совершать медленные движения
- 10. Механизмы быстрого сна Нейроны ствола, вовлеченные в регуляцию быстрого сна, можно разделить на 3 группы: REM-on
- 11. Переключение между фазами сна REM-off нейроны серотонин норадреналин REM-on нейроны АцХ глутамат ГАМК-эргические нейроны ВЛПЯ, среднего
- 12. Фаза быстрого сна Характеризуется быстрыми движениями глаз, поэтому ее называют сон с быстрыми движениями глаз, или
- 13. Цикл сна Таким образом, вся эта последовательность называется циклом. Первый цикл имеет длительность 90-100 мин. Затем
- 14. Механизмы циркадианной ритмики (1) Циркадианные ритмы – физиологические и психические явления, возникающие с периодом около суток.
- 15. Механизмы циркадианной ритмики (2) Обычно цикл реакций у большинства людей составляет 25 ч ? нужна «ежедневная»
- 16. Механизмы циркадианной ритмики (3) 2. Мелатонин СХЯ и эпифиз — две половины главных «биологических часов», находящиеся
- 17. Восстановление общего и электролитного гомеостаза мозговой ткани (NREM-сон) Значение сна Оптимизация управления внутренними органами (NREM-сон) Переработка
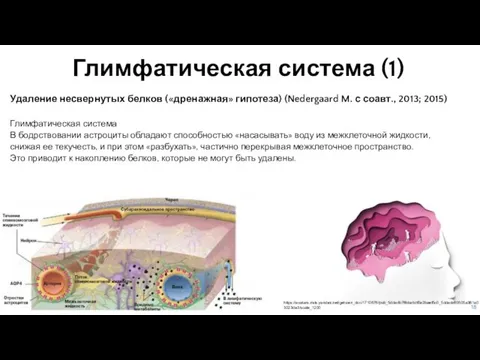
- 18. Глимфатическая система (1) Удаление несвернутых белков («дренажная» гипотеза) (Nedergaard M. с соавт., 2013; 2015) Глимфатическая система
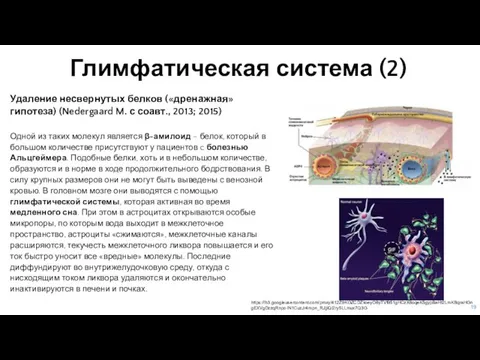
- 19. Глимфатическая система (2) Удаление несвернутых белков («дренажная» гипотеза) (Nedergaard M. с соавт., 2013; 2015) Одной из
- 20. Ковальзон В.М. Основы сомнологии. Физиология и нейрохимия цикла «бодрствование-сон». М.: БИНОМ. Лаборатория знаний; 2012. Петров А.М.,
- 22. Скачать презентацию
Слайд 3Сон – это особое генетически детерминированное состояние организма гомеотермных (теплокровных) животных, характеризующееся
Сон – это особое генетически детерминированное состояние организма гомеотермных (теплокровных) животных, характеризующееся

Слайд 4Цикл «бодрствование – сон»
В регуляции цикла «бодрствование-сон» на системном уровне принимают
Цикл «бодрствование – сон»
В регуляции цикла «бодрствование-сон» на системном уровне принимают

Слайд 5Механизмы поддержания бодрствования (1)
На уровне ромбовидного мозга:
нейроны синего пятна (норадреналин)
нейроны педункулопонтинной и
Механизмы поддержания бодрствования (1)
На уровне ромбовидного мозга:
нейроны синего пятна (норадреналин)
нейроны педункулопонтинной и

Слайд 6Под воздействием восходящей активации нейроны коры переходят в состояние так называемой «тонической
Под воздействием восходящей активации нейроны коры переходят в состояние так называемой «тонической

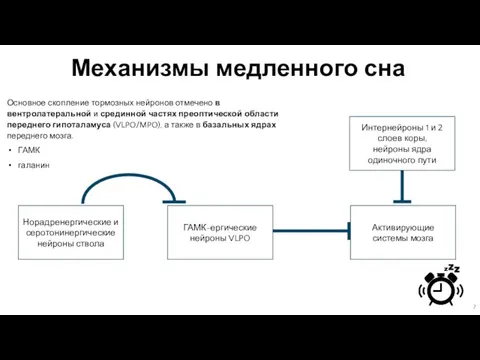
Слайд 7Механизмы медленного сна
Основное скопление тормозных нейронов отмечено в вентролатеральной и срединной частях
Механизмы медленного сна
Основное скопление тормозных нейронов отмечено в вентролатеральной и срединной частях
Слайд 8Роль аденозина
Одна из гипотез связывает причину медленного сна с постепенным накоплением в
Роль аденозина
Одна из гипотез связывает причину медленного сна с постепенным накоплением в

Слайд 9Стадии медленного сна
1 стадия (дремота):
5-10 мин
снижается мышечная активность
глаза могут совершать медленные движения
тета-ритм
Стадии медленного сна
1 стадия (дремота):
5-10 мин
снижается мышечная активность
глаза могут совершать медленные движения
тета-ритм

Слайд 10Механизмы быстрого сна
Нейроны ствола, вовлеченные в регуляцию быстрого сна, можно разделить на
Механизмы быстрого сна
Нейроны ствола, вовлеченные в регуляцию быстрого сна, можно разделить на

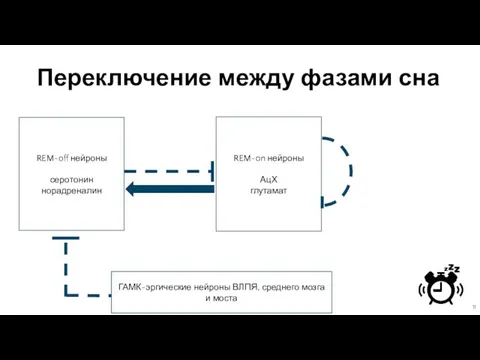
Слайд 11Переключение между фазами сна
REM-off нейроны
серотонин
норадреналин
REM-on нейроны
АцХ
глутамат
ГАМК-эргические нейроны ВЛПЯ, среднего мозга и моста
Переключение между фазами сна
REM-off нейроны
серотонин
норадреналин
REM-on нейроны
АцХ
глутамат
ГАМК-эргические нейроны ВЛПЯ, среднего мозга и моста
Слайд 12Фаза быстрого сна
Характеризуется быстрыми движениями глаз, поэтому ее называют сон с быстрыми
Фаза быстрого сна
Характеризуется быстрыми движениями глаз, поэтому ее называют сон с быстрыми

Слайд 13Цикл сна
Таким образом, вся эта последовательность называется циклом.
Первый цикл имеет длительность
Цикл сна
Таким образом, вся эта последовательность называется циклом.
Первый цикл имеет длительность

Слайд 14Механизмы циркадианной ритмики (1)
Циркадианные ритмы – физиологические и психические явления, возникающие
Механизмы циркадианной ритмики (1)
Циркадианные ритмы – физиологические и психические явления, возникающие

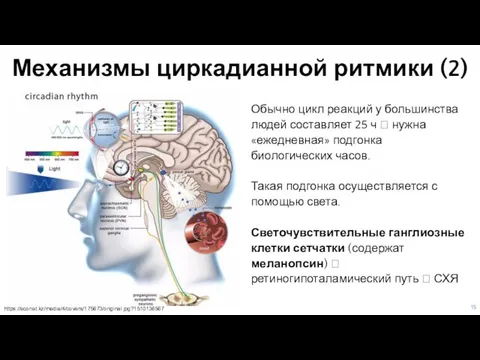
Слайд 15Механизмы циркадианной ритмики (2)
Обычно цикл реакций у большинства людей составляет 25
Механизмы циркадианной ритмики (2)
Обычно цикл реакций у большинства людей составляет 25
Слайд 16Механизмы циркадианной ритмики (3)
2. Мелатонин
СХЯ и эпифиз — две половины главных
Механизмы циркадианной ритмики (3)
2. Мелатонин
СХЯ и эпифиз — две половины главных

Слайд 17Восстановление общего и электролитного гомеостаза мозговой ткани (NREM-сон)
Значение сна
Оптимизация управления внутренними органами
Восстановление общего и электролитного гомеостаза мозговой ткани (NREM-сон)
Значение сна
Оптимизация управления внутренними органами

Слайд 18Глимфатическая система (1)
Удаление несвернутых белков («дренажная» гипотеза) (Nedergaard M. с соавт., 2013;
Глимфатическая система (1)
Удаление несвернутых белков («дренажная» гипотеза) (Nedergaard M. с соавт., 2013;
Слайд 19Глимфатическая система (2)
Удаление несвернутых белков («дренажная» гипотеза) (Nedergaard M. с соавт., 2013;
Глимфатическая система (2)
Удаление несвернутых белков («дренажная» гипотеза) (Nedergaard M. с соавт., 2013;
Слайд 20Ковальзон В.М. Основы сомнологии. Физиология и нейрохимия цикла «бодрствование-сон». М.: БИНОМ. Лаборатория
Ковальзон В.М. Основы сомнологии. Физиология и нейрохимия цикла «бодрствование-сон». М.: БИНОМ. Лаборатория

 Шок. Острая кровопотеря
Шок. Острая кровопотеря Клинический случай
Клинический случай Дифтерийная инфекция
Дифтерийная инфекция СРС: DOTS программа или этиотропное лечение
СРС: DOTS программа или этиотропное лечение Корь
Корь Особенности организма детей среднего и старшего возраста
Особенности организма детей среднего и старшего возраста Клиническая фармакология стероидных и нестероидных противовоспалительных препаратов
Клиническая фармакология стероидных и нестероидных противовоспалительных препаратов Эшерихиозы
Эшерихиозы Синдром Шарпа
Синдром Шарпа Коллагеноздар. Эритематоз. Этиологиясы, патогенезі, бейімділік факторлары,
Коллагеноздар. Эритематоз. Этиологиясы, патогенезі, бейімділік факторлары, Імунна відповідь у нормі та при її порушення при найбільш поширених патологічних станах
Імунна відповідь у нормі та при її порушення при найбільш поширених патологічних станах Лаборатория остаточных элементов. Химия тела
Лаборатория остаточных элементов. Химия тела Контроль вирусных вакцин на биофабрике
Контроль вирусных вакцин на биофабрике Эписклеральное секторальное пломбирование
Эписклеральное секторальное пломбирование История развития психопатологии в России и зарубежных странах
История развития психопатологии в России и зарубежных странах Желтухи при полицитемиях у новорожденных
Желтухи при полицитемиях у новорожденных №3, посев
№3, посев Травмы. Раны. Кровотечения. Школа медицинских волонтеров
Травмы. Раны. Кровотечения. Школа медицинских волонтеров file-4014 (1)
file-4014 (1) Специфическая аллергологическая диагностика. Вариант 1
Специфическая аллергологическая диагностика. Вариант 1 Глоссарий
Глоссарий Predominantly unconjugated hyperbilirubinemia
Predominantly unconjugated hyperbilirubinemia ТЕРМИНАЛЬНЫЕ СОСТОЯНИЯ
ТЕРМИНАЛЬНЫЕ СОСТОЯНИЯ Ерте жастағы балалардағы респираторлық ауруларды диагностикалаумен емдеудің заманауи аспектілері. Бронхтың обструктивті
Ерте жастағы балалардағы респираторлық ауруларды диагностикалаумен емдеудің заманауи аспектілері. Бронхтың обструктивті ОҚО бүйрек трансплантациясы
ОҚО бүйрек трансплантациясы Понятие и виды психических процессов
Понятие и виды психических процессов Созылмалы бүйрек ауруларының үдеу механизмі
Созылмалы бүйрек ауруларының үдеу механизмі Вакцинопрофилактика. Новокузнецкий центр общественного здоровья и медицинской профилактики
Вакцинопрофилактика. Новокузнецкий центр общественного здоровья и медицинской профилактики