- Органы иммунной системы человека

Содержание
- 2. Красный костный мозг Это вещество, которое заполняет всё внутреннее пространство костной губчатой ткани. Самая большая концентрация
- 3. Основные функции селезёнки: высвобождение накопленной крови, повышение общего кровоснабжения организма и обогащение тканей тела кислородом; разрушение
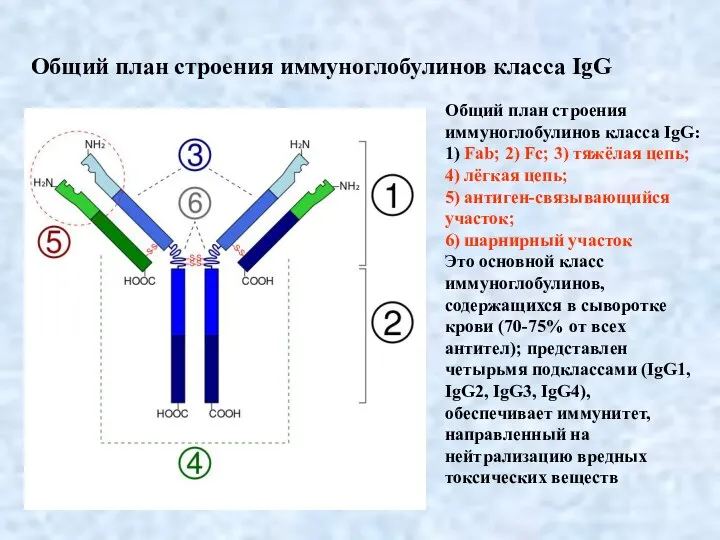
- 4. Общий план строения иммуноглобулинов класса IgG: 1) Fab; 2) Fc; 3) тяжёлая цепь; 4) лёгкая цепь;
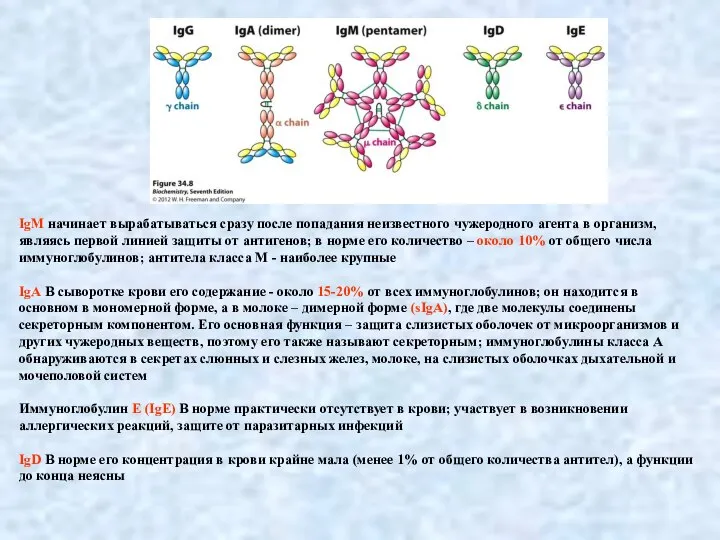
- 5. IgM начинает вырабатываться сразу после попадания неизвестного чужеродного агента в организм, являясь первой линией защиты от
- 6. ГЕНЕТИКА ИММУНОГЛОБУЛИНОВ В биологии существовует принцип «один ген — одна полипептидная цепь. Однако в случае Ig
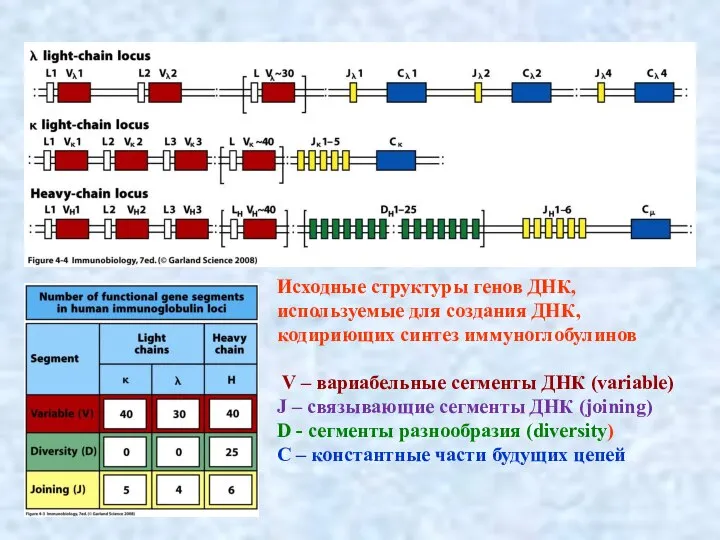
- 7. Исходные структуры генов ДНК, используемые для создания ДНК, кодириющих синтез иммуноглобулинов V – вариабельные сегменты ДНК
- 8. Перестройка ДНК из разных фрагментов V и J сегментов Кодирующая ДНК для каждой из легких цепей
- 9. Перестройка зародышевой ДНК для тяжелых цепей происходит из разных фрагментов V, J и D сегментов Кодирующая
- 10. При синтезе антител разных классов происходит переключение к набору фрагментов из разных частей константных частей зародышевых
- 11. Переключение синтеза генов, кодирующих иммуноглобулины одного класса на другой происходит с помощью рекомбинации ДНК Например, сначала
- 12. Переключение синтеза генов, кодирующих иммуноглобулины одного класса на другой происходит с помощью рекомбинации ДНК Например, сначала
- 13. В результате рекомбинации в непрерывную цепь ДНК соединяется по одному сегменту V-, D и J-областей -
- 14. Теоретически В результате рекомбинаций число вариантов антител по одной специфичности к одному антигену исходя из правил
- 15. Не все варианты рекомбинаций абсолютно правильные – эффективные Под неточностью формирования VDJ понимают тот факт, что
- 16. Одним из четких показателей повышенной техногенной нагрузки у различных групп населения является повышенный процент индивидов, проявляющих
- 17. В норме иммунная система человека четко различает “свое” и “чужое”. В крови здоровых людей отсутствуют антитела
- 18. Фундаментальным и наиболее исследованным свойством иммуноглобулинов является их способность связывать и нейтрализовать самые разнообразные по структуре
- 19. В настоящее время показано существование двух путей наработки антител ферментов. Первый путь – наработка антител к
- 20. Еще одним путем генерации индуцированных абзимов является аутоиммунизация организма различными ферментами и наработка антител против активных
- 21. К настоящему моменту открыты природные абзимы, гидролизующие: 1. Белки 2. ДНК 3. РНК 4. Полисахариды 5.
- 22. К настоящему моменту показано, что ДНК-гидролизующие абзимы отсутствуют у здоровых доноров, но есть в крови пациентов
- 23. а) Совпадение положения пиков IgG и ДНКазной активности при гель-фильтрации в кислом буфере г) Совпадение положения
- 24. Показано, что анализ относительной активности антител в гидролизе ДНК может быть использован для оценки глубины аутоиммунных
- 25. Эффект лечения больных аутоиммунным тиреоидитом плаквинилом
- 26. Относительная активность и субстрантная специфичность РНК-гидролизующих антитед очень сильно зависит от типа аутоиммунного заболевания Гидролиз тРНК
- 27. Впервые показано, что абзимы крови больных рассеянным склерозом специфично гидролизуют основный белок миелина – белково-липидной оболочки
- 28. а) Гидролиз ОБМ б) Адсорбция АТ человека антителаами мышей против IgG и элюция кислым буфером в)
- 29. На примере антител, гидролизующих основной белок миелина, нами впервые показано, что антитела с протеолитической активностью могут
- 30. Зависимые и независимые от ионов металлов антитела, гидролизующие основной белок миелина обнаружены в крови пациентов с:
- 31. В крови больных ВИЧ-инфецированных больных кроме ДНК-. РНК- гидролизующих, обнаружены антитела гидролизующие вирусные обратную транскриптазу и
- 32. Как указывалось ранее, теоретически количество антител против одного антигена может быть очень большим – до одного
- 33. Одним из путей разделения разных абзимов является аффинная хроматография на сорбентах с иммобилизованными субстратами. Например, аффинная
- 34. . Множество разных рН оптимумов в гидролизе ДНК антителами из крови разных больных СКВ Такое же
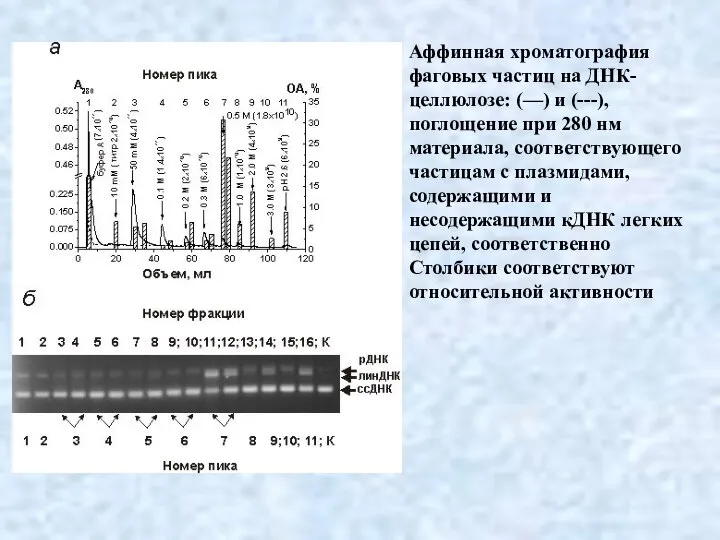
- 35. Аффинная хроматография фаговых частиц на ДНК-целлюлозе: (—) и (---), поглощение при 280 нм материала, соответствующего частицам
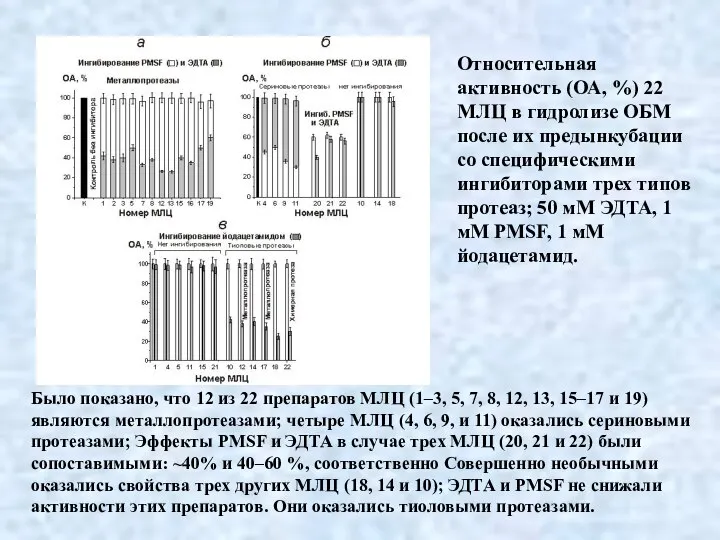
- 36. Относительная активность (ОА, %) 22 МЛЦ в гидролизе ОБМ после их предынкубации со специфическими ингибиторами трех
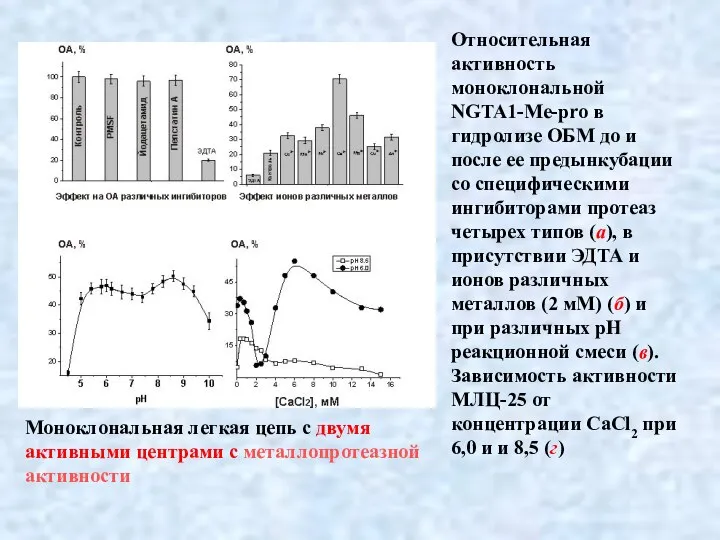
- 37. Относительная активность моноклональной NGTA1-Me-pro в гидролизе ОБМ до и после ее предынкубации со специфическими ингибиторами протеаз
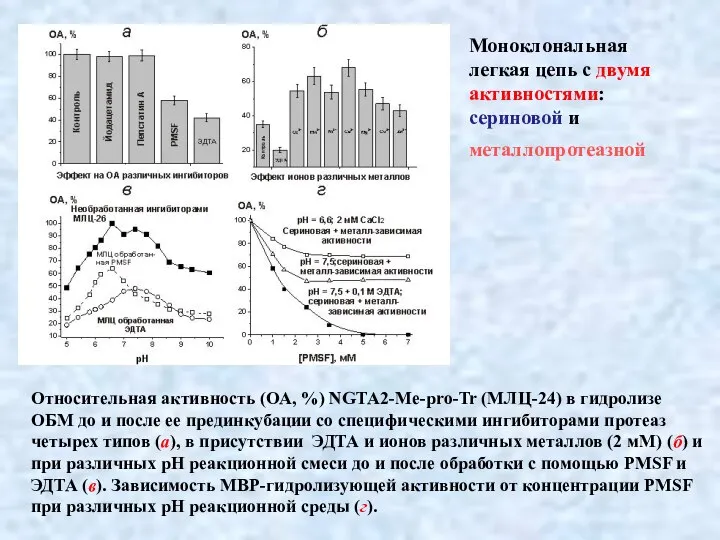
- 38. Относительная активность (ОА, %) NGTA2-Me-pro-Tr (МЛЦ-24) в гидролизе ОБМ до и после ее прединкубации со специфическими
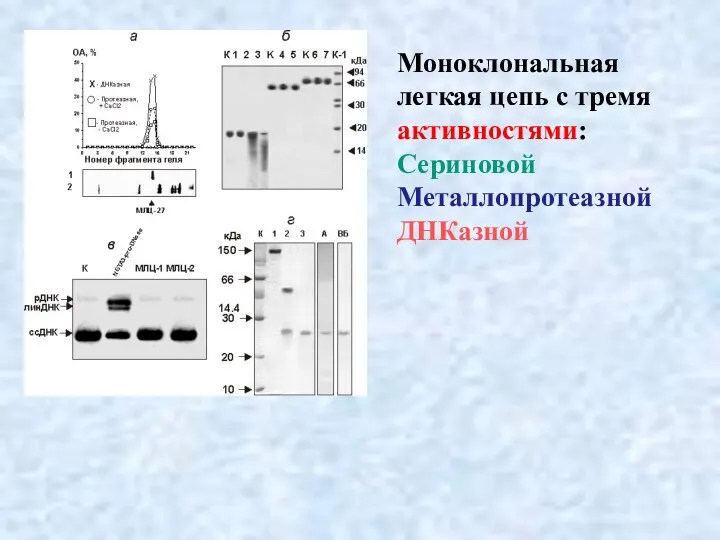
- 39. Моноклональная легкая цепь с тремя активностями: Сериновой Металлопротеазной ДНКазной
- 40. Современная теория кроветворения, основанная на унитарной теории отечественного гистолога А.А. Максимова, различает шесть классов кроветворных клеток.
- 41. 4) Моноцитарно-макрофагальному. На территории костного мозга дифференцировка в данном направлении завершается образованием моноцитов, мигрирующих в кровь;
- 43. Скачать презентацию
Слайд 2Красный костный мозг
Это вещество, которое заполняет всё внутреннее пространство костной губчатой ткани.
Красный костный мозг
Это вещество, которое заполняет всё внутреннее пространство костной губчатой ткани.
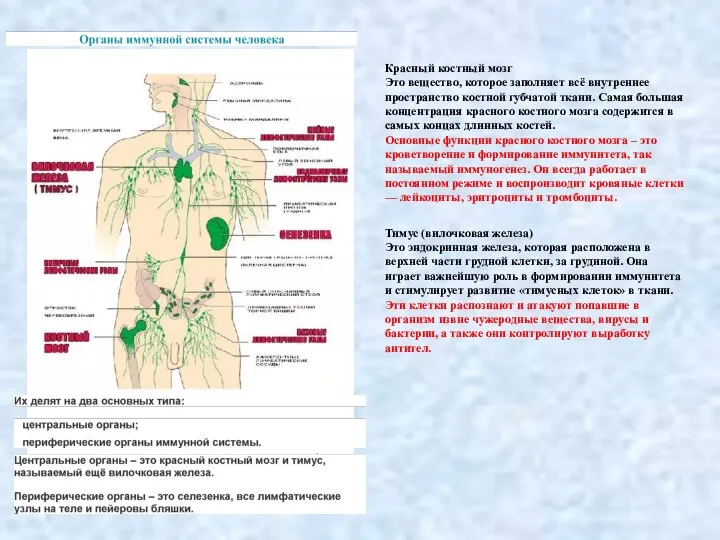
Слайд 3Основные функции селезёнки:
высвобождение накопленной крови, повышение общего кровоснабжения организма и обогащение тканей
Основные функции селезёнки:
высвобождение накопленной крови, повышение общего кровоснабжения организма и обогащение тканей

Слайд 4Общий план строения иммуноглобулинов класса IgG:
1) Fab; 2) Fc; 3) тяжёлая цепь;
Общий план строения иммуноглобулинов класса IgG:
1) Fab; 2) Fc; 3) тяжёлая цепь;
Слайд 5IgM начинает вырабатываться сразу после попадания неизвестного чужеродного агента в организм, являясь
IgM начинает вырабатываться сразу после попадания неизвестного чужеродного агента в организм, являясь
Слайд 6ГЕНЕТИКА ИММУНОГЛОБУЛИНОВ
В биологии существовует принцип «один ген — одна полипептидная цепь. Однако
ГЕНЕТИКА ИММУНОГЛОБУЛИНОВ
В биологии существовует принцип «один ген — одна полипептидная цепь. Однако

Слайд 7Исходные структуры генов ДНК, используемые для создания ДНК, кодириющих синтез иммуноглобулинов
Исходные структуры генов ДНК, используемые для создания ДНК, кодириющих синтез иммуноглобулинов
Слайд 8Перестройка ДНК из разных фрагментов V и J сегментов
Кодирующая ДНК для
Перестройка ДНК из разных фрагментов V и J сегментов
Кодирующая ДНК для
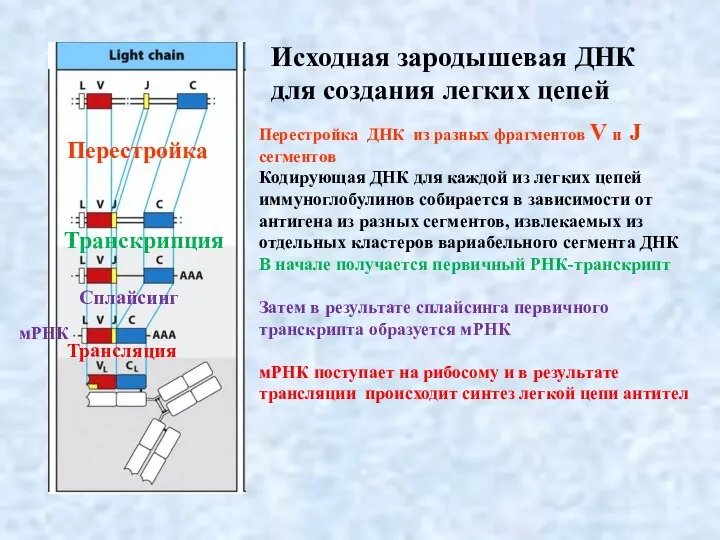
Слайд 9Перестройка зародышевой ДНК для тяжелых цепей происходит из разных фрагментов V, J
Перестройка зародышевой ДНК для тяжелых цепей происходит из разных фрагментов V, J
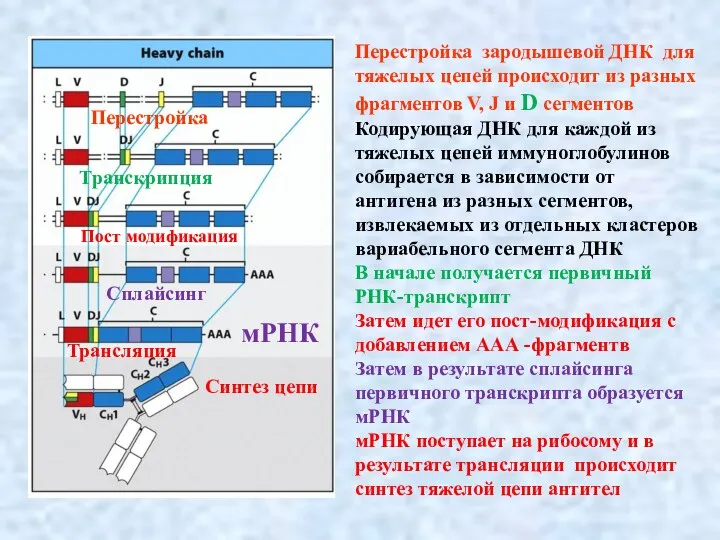
Слайд 10При синтезе антител разных классов происходит переключение к набору фрагментов из разных
При синтезе антител разных классов происходит переключение к набору фрагментов из разных
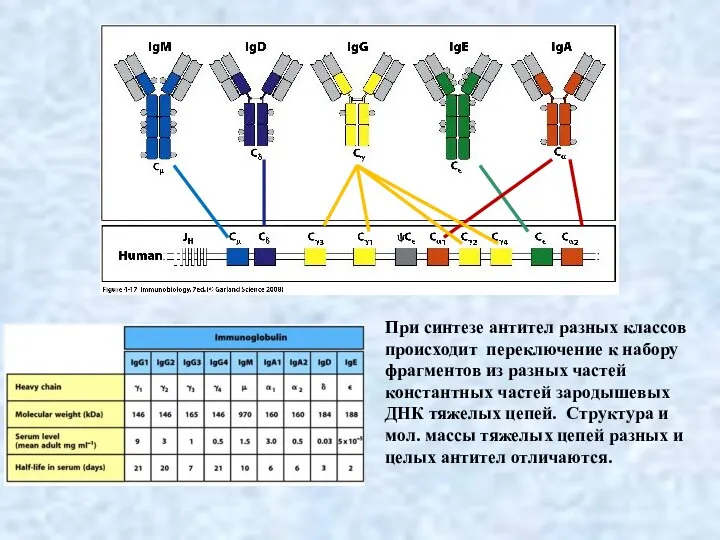
Слайд 11Переключение синтеза генов, кодирующих иммуноглобулины одного класса на другой происходит с помощью
Переключение синтеза генов, кодирующих иммуноглобулины одного класса на другой происходит с помощью
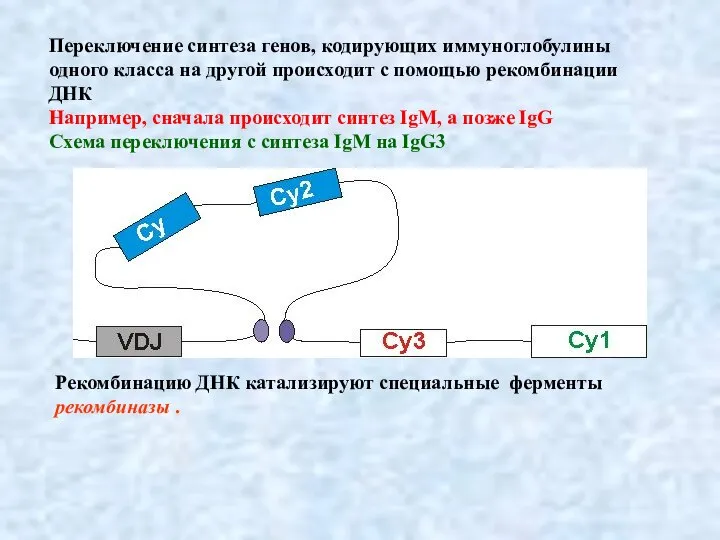
Слайд 12Переключение синтеза генов, кодирующих иммуноглобулины одного класса на другой происходит с помощью
Переключение синтеза генов, кодирующих иммуноглобулины одного класса на другой происходит с помощью
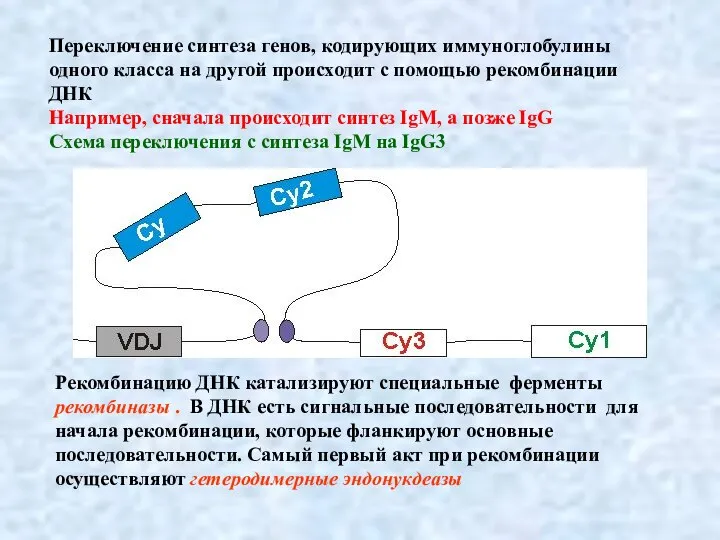
Слайд 13В результате рекомбинации в непрерывную цепь ДНК соединяется
по одному сегменту V-,
В результате рекомбинации в непрерывную цепь ДНК соединяется
по одному сегменту V-,

Слайд 14Теоретически В результате рекомбинаций число
вариантов антител по одной специфичности к одному
Теоретически В результате рекомбинаций число
вариантов антител по одной специфичности к одному

Слайд 15Не все варианты рекомбинаций абсолютно
правильные – эффективные
Под неточностью формирования VDJ понимают
Не все варианты рекомбинаций абсолютно
правильные – эффективные
Под неточностью формирования VDJ понимают

Слайд 16Одним из четких показателей повышенной техногенной нагрузки у различных групп населения является
Одним из четких показателей повышенной техногенной нагрузки у различных групп населения является

Слайд 17В норме иммунная система человека четко различает “свое” и “чужое”. В крови
В норме иммунная система человека четко различает “свое” и “чужое”. В крови

Слайд 18Фундаментальным и наиболее исследованным свойством иммуноглобулинов является их способность связывать и нейтрализовать
Фундаментальным и наиболее исследованным свойством иммуноглобулинов является их способность связывать и нейтрализовать

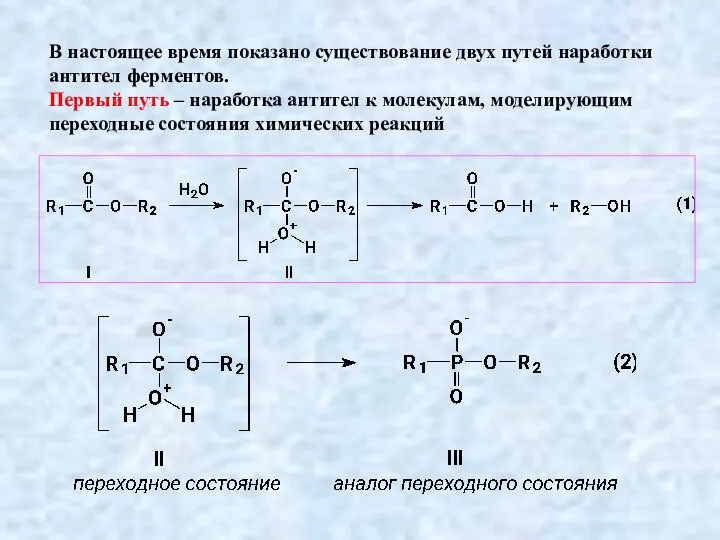
Слайд 19В настоящее время показано существование двух путей наработки антител ферментов.
Первый путь –
В настоящее время показано существование двух путей наработки антител ферментов.
Первый путь –
Слайд 20
Еще одним путем генерации индуцированных абзимов является аутоиммунизация организма различными ферментами и
Еще одним путем генерации индуцированных абзимов является аутоиммунизация организма различными ферментами и

Слайд 21К настоящему моменту открыты природные абзимы, гидролизующие:
1. Белки
2. ДНК
3. РНК
4. Полисахариды
К настоящему моменту открыты природные абзимы, гидролизующие:
1. Белки
2. ДНК
3. РНК
4. Полисахариды

Слайд 22К настоящему моменту показано, что ДНК-гидролизующие абзимы отсутствуют у здоровых доноров, но
К настоящему моменту показано, что ДНК-гидролизующие абзимы отсутствуют у здоровых доноров, но

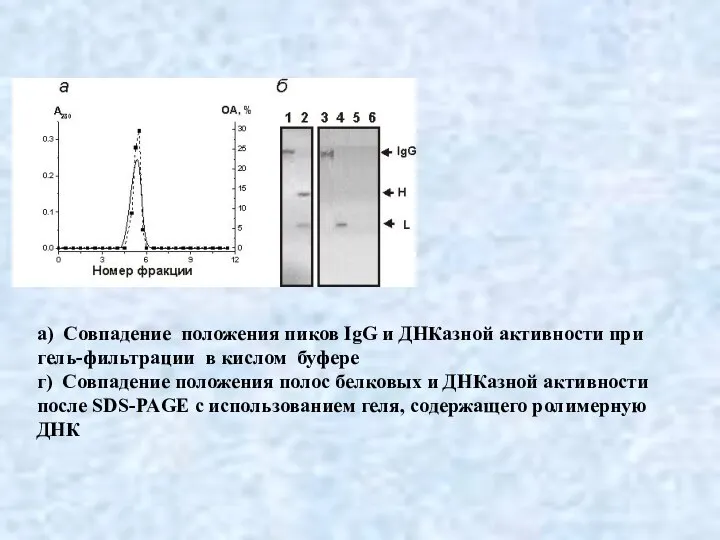
Слайд 23а) Совпадение положения пиков IgG и ДНКазной активности при
гель-фильтрации в кислом буфере
а) Совпадение положения пиков IgG и ДНКазной активности при
гель-фильтрации в кислом буфере
Слайд 24Показано, что анализ относительной активности антител в гидролизе ДНК может быть использован
Показано, что анализ относительной активности антител в гидролизе ДНК может быть использован

Слайд 25Эффект лечения больных
аутоиммунным тиреоидитом плаквинилом
Эффект лечения больных
аутоиммунным тиреоидитом плаквинилом

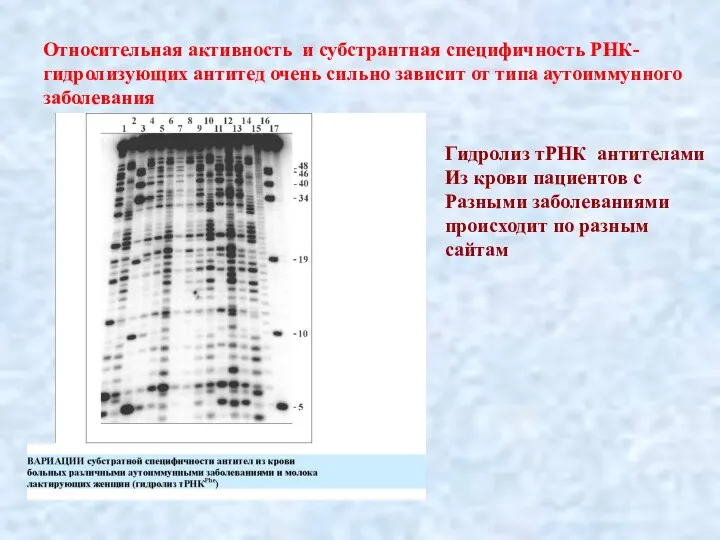
Слайд 26Относительная активность и субстрантная специфичность РНК-гидролизующих антитед очень сильно зависит от типа
Относительная активность и субстрантная специфичность РНК-гидролизующих антитед очень сильно зависит от типа
Слайд 27Впервые показано, что абзимы крови больных рассеянным склерозом специфично гидролизуют основный белок
Впервые показано, что абзимы крови больных рассеянным склерозом специфично гидролизуют основный белок

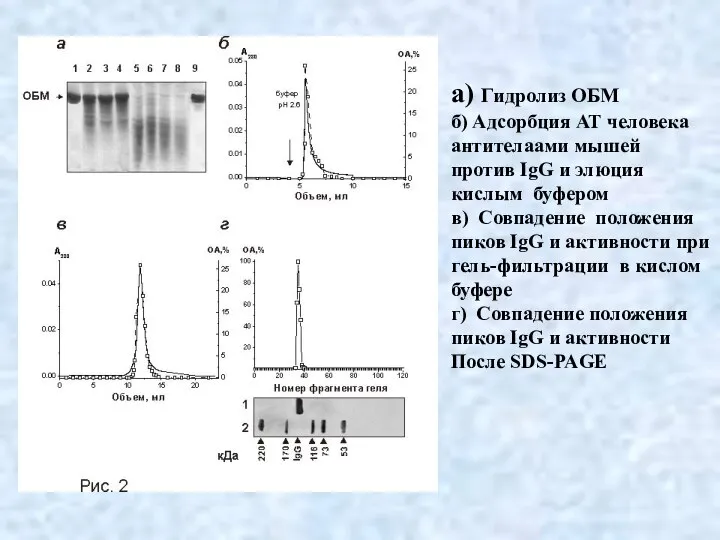
Слайд 28а) Гидролиз ОБМ
б) Адсорбция АТ человека
антителаами мышей
против IgG и элюция
кислым
а) Гидролиз ОБМ
б) Адсорбция АТ человека
антителаами мышей
против IgG и элюция
кислым
Слайд 29На примере антител, гидролизующих основной белок миелина, нами впервые показано, что антитела
На примере антител, гидролизующих основной белок миелина, нами впервые показано, что антитела

Слайд 30Зависимые и независимые от ионов металлов антитела, гидролизующие основной белок миелина обнаружены
Зависимые и независимые от ионов металлов антитела, гидролизующие основной белок миелина обнаружены

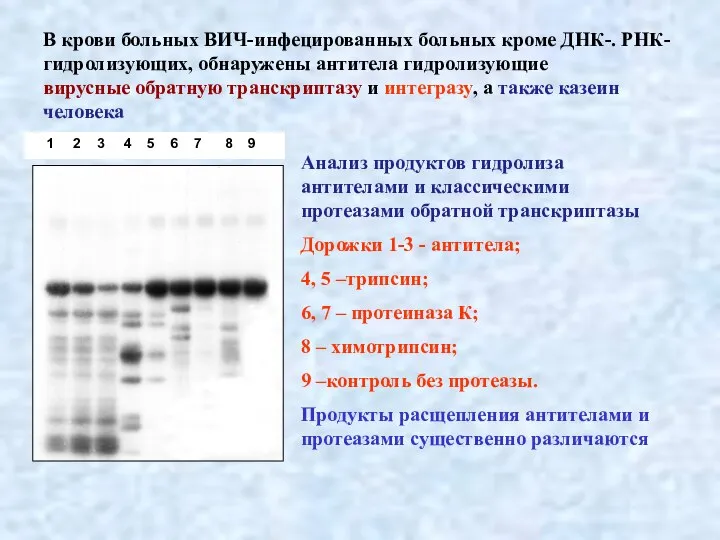
Слайд 31В крови больных ВИЧ-инфецированных больных кроме ДНК-. РНК- гидролизующих, обнаружены антитела гидролизующие
В крови больных ВИЧ-инфецированных больных кроме ДНК-. РНК- гидролизующих, обнаружены антитела гидролизующие
Слайд 32Как указывалось ранее, теоретически количество антител против одного антигена может быть очень
Как указывалось ранее, теоретически количество антител против одного антигена может быть очень

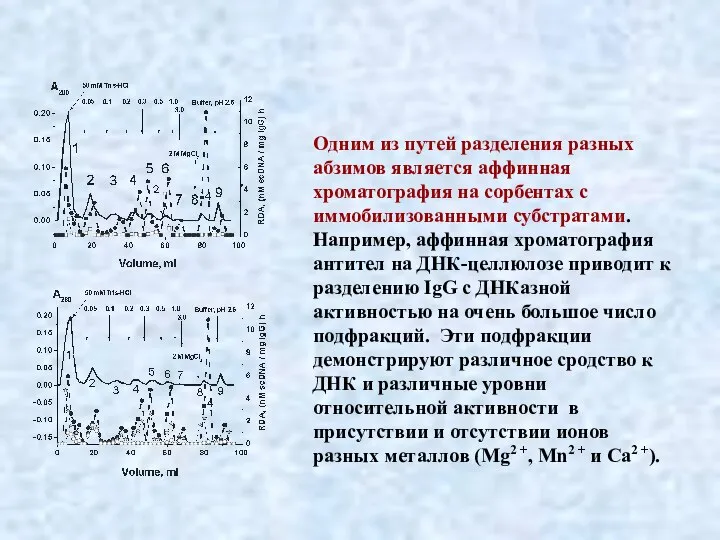
Слайд 33Одним из путей разделения разных абзимов является аффинная хроматография на сорбентах с
Одним из путей разделения разных абзимов является аффинная хроматография на сорбентах с
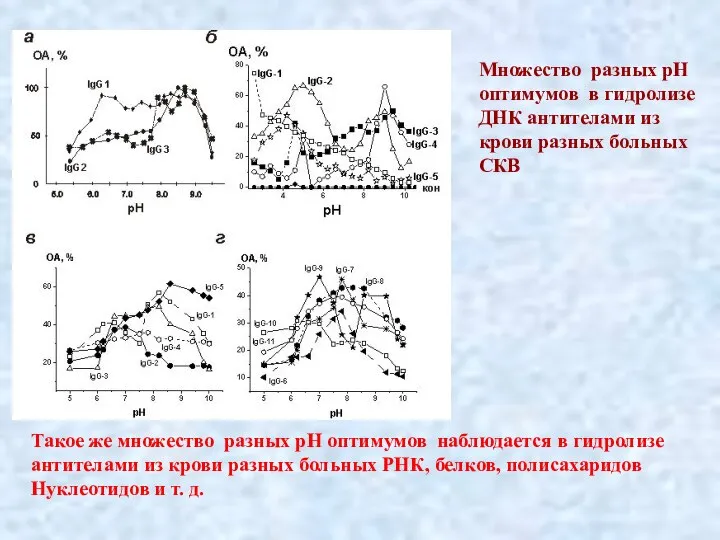
Слайд 34.
Множество разных рН
оптимумов в гидролизе
ДНК антителами из
крови разных больных
СКВ
Такое же
.
Множество разных рН
оптимумов в гидролизе
ДНК антителами из
крови разных больных
СКВ
Такое же
Слайд 35Аффинная хроматография фаговых частиц на ДНК-целлюлозе: (—) и (---), поглощение при 280
Аффинная хроматография фаговых частиц на ДНК-целлюлозе: (—) и (---), поглощение при 280
Слайд 36Относительная активность (ОА, %) 22 МЛЦ в гидролизе ОБМ после их предынкубации
Относительная активность (ОА, %) 22 МЛЦ в гидролизе ОБМ после их предынкубации
Слайд 37Относительная активность моноклональной NGTA1-Me-pro в гидролизе ОБМ до и после ее предынкубации
Относительная активность моноклональной NGTA1-Me-pro в гидролизе ОБМ до и после ее предынкубации
Слайд 38Относительная активность (ОА, %) NGTA2-Me-pro-Tr (МЛЦ-24) в гидролизе ОБМ до и после
Относительная активность (ОА, %) NGTA2-Me-pro-Tr (МЛЦ-24) в гидролизе ОБМ до и после
Слайд 39Моноклональная
легкая цепь с тремя активностями:
Сериновой
Металлопротеазной
ДНКазной
Моноклональная
легкая цепь с тремя активностями:
Сериновой
Металлопротеазной
ДНКазной
Слайд 40Современная теория кроветворения, основанная на унитарной теории отечественного гистолога А.А. Максимова, различает
Современная теория кроветворения, основанная на унитарной теории отечественного гистолога А.А. Максимова, различает

Слайд 414) Моноцитарно-макрофагальному. На территории костного мозга дифференцировка в данном направлении завершается образованием
4) Моноцитарно-макрофагальному. На территории костного мозга дифференцировка в данном направлении завершается образованием

 Фармакологияның даму тарихы
Фармакологияның даму тарихы Лабораторная диагностика паразитов
Лабораторная диагностика паразитов Паразитарные заболевания, вызываемые паразитами животной природы
Паразитарные заболевания, вызываемые паразитами животной природы Ведение регистров пациентов с новой коронавирусной инфекцией Covid-19 и внебольничними пневмониями
Ведение регистров пациентов с новой коронавирусной инфекцией Covid-19 и внебольничними пневмониями Сепсис. Лечение сепсиса
Сепсис. Лечение сепсиса Транспозиция магистральных сосудов
Транспозиция магистральных сосудов Приемы временной остановки наружного кровотечения
Приемы временной остановки наружного кровотечения Легочная эмфизема семейная у детей
Легочная эмфизема семейная у детей Громкие вспышки сибирской язвы
Громкие вспышки сибирской язвы Патологиялық материалды өңдеу тәртібі. Зерттеуге вирус бар материалды дайындау
Патологиялық материалды өңдеу тәртібі. Зерттеуге вирус бар материалды дайындау Материалы к итоговой аттестации интернов-терапевтов 2018 год
Материалы к итоговой аттестации интернов-терапевтов 2018 год Лабораторная диагностика сифилиса и цитомегаловирусной инфекции
Лабораторная диагностика сифилиса и цитомегаловирусной инфекции Актуальные вопросы организации медицинской помощи пациентам с новой коронавирусной инфекцией
Актуальные вопросы организации медицинской помощи пациентам с новой коронавирусной инфекцией Пищеварение в ротовой полости. Зубы
Пищеварение в ротовой полости. Зубы Уход за новорожденным в стационаре
Уход за новорожденным в стационаре Лекарственные растения
Лекарственные растения Гнойно-воспалительные заболевания женских половых органов
Гнойно-воспалительные заболевания женских половых органов Психолого-присихиатрические признаки эпилепсии
Психолого-присихиатрические признаки эпилепсии Респираторный микоплазмоз
Респираторный микоплазмоз Увеиты при склеродермии
Увеиты при склеродермии Примеры эффективности применения БАК у детей в клинике
Примеры эффективности применения БАК у детей в клинике Правильное питание - основа здоровья
Правильное питание - основа здоровья Антигиперурекимические средства
Антигиперурекимические средства Офтальмохирургия
Офтальмохирургия Культура здоровья
Культура здоровья Заболевание холера
Заболевание холера Первая помощь пострадавшему во время занятий физическими упражнениями при обмороке
Первая помощь пострадавшему во время занятий физическими упражнениями при обмороке Травма. Диагностика. Лечение. Синдром длительного сдавливания
Травма. Диагностика. Лечение. Синдром длительного сдавливания