- Яичниковый цикл

Содержание
- 2. Фазы менструального цикла : 1-я (фолликулярная) фаза цикла определяется ростом и созреванием фолликула и яйцеклетки в
- 3. Уровни регуляции Первый (высший) уровень- кора головного мозга и экстрагипоталамические церебральные структуры (лимбическая система, гиппокамп, миндалевидное
- 4. Первый уровень регуляции : ЦНС Адекватное состояние ЦНС обеспечивает нормальное функционирование всех нижележащих звеньев репродуктивной системы.
- 5. Второй уровень регуляции: гипоталамус Гипоталамус участвует в регуляции полового поведения, осуществляет контроль за вегетососудистыми реакциями, температурой
- 6. Установлено, что люлиберин, или рилизинг-гормон лютеинизирующего гормона (РГЛГ) и его синтетические аналоги стимулируют выделение гонадотрофами не
- 7. Третьим уровнем регуляции- гипофиз Гипофиз состоит из передней, задней и промежуточной (средней) доли. Передняя доля (аденогипофиз)
- 8. Четвертому уровню регуляции- яичники Функции яичника : Генеративная функция (рост и созревание фолликулов, созревание яйцеклетки) Гормональная
- 9. Выделяют четыре типа фолликула: Примордиальный фолликул состоит из незрелой яйцеклетки (ооцита) в профазе 2-го мейотического деления,
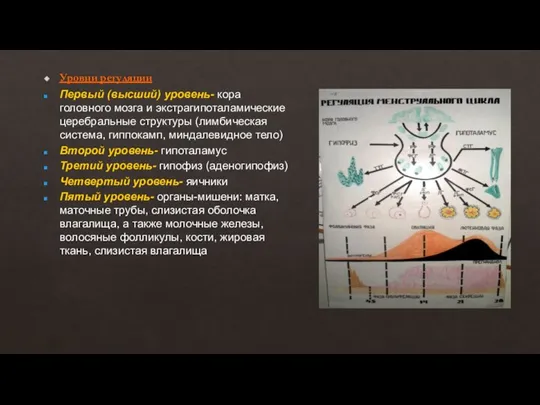
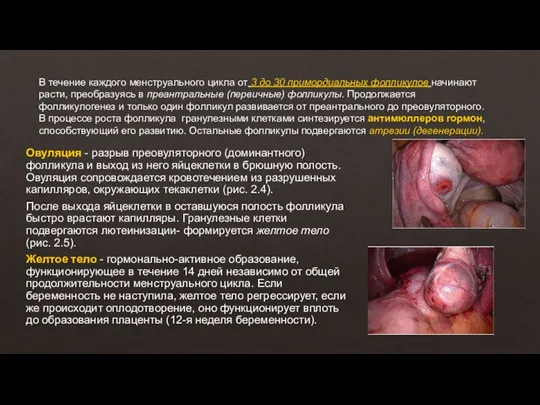
- 10. В течение каждого менструального цикла от 3 до 30 примордиальных фолликулов начинают расти, преобразуясь в преантральные
- 11. Гормональная функция яичников Рост, созревание фолликулов в яичниках и образование желтого тела сопровождаются продукцией половых стероидных
- 12. Цикличность деятельности репродуктивной системы определяется принципами прямой и обратной связи, которая обеспечивается специфическими рецепторами к гормонам
- 13. Яичниковый цикл Фолликулярная фаза: Пульсирующая секреция и выделение ГнРГ приводят к высвобождению ФСГ и ЛГ из
- 14. Лютеиновая фаза: После овуляции уровень ЛГ снижается по отношению к «овуляторному пику». Данное количество ЛГ стимулирует:
- 15. Пятый уровень регуляции органы-мишени: матка, маточные трубы, слизистая оболочка влагалища, а также молочные железы, волосяные фолликулы,
- 16. В эндометрии в течение менструального цикла происходят следующие изменения: фаза десквамация и отторжение функционального слоя, фаза
- 17. Фаза секреции (лютеиновая фаза в яичниках) связана с активностью желтого тела, длится 14±1 день. В этот
- 18. Менструация включает десквамацию, отторжение и регенерацию функционального слоя эндометрия. В связи с регрессом желтого тела и
- 19. Регенерация эндометрия начинается сразу после отторжения некротизированного функционального слоя. Основой для регенерации являются эпителиальные клетки стромы
- 21. Скачать презентацию

Слайд 3Уровни регуляции
Первый (высший) уровень- кора головного мозга и экстрагипоталамические церебральные структуры (лимбическая
Уровни регуляции
Первый (высший) уровень- кора головного мозга и экстрагипоталамические церебральные структуры (лимбическая
Слайд 4Первый уровень регуляции : ЦНС
Адекватное состояние ЦНС обеспечивает нормальное функционирование всех нижележащих
Первый уровень регуляции : ЦНС
Адекватное состояние ЦНС обеспечивает нормальное функционирование всех нижележащих

Слайд 5Второй уровень регуляции: гипоталамус
Гипоталамус участвует в регуляции полового поведения, осуществляет контроль за
Второй уровень регуляции: гипоталамус
Гипоталамус участвует в регуляции полового поведения, осуществляет контроль за

Слайд 6Установлено, что люлиберин, или рилизинг-гормон лютеинизирующего гормона (РГЛГ) и его синтетические аналоги
Установлено, что люлиберин, или рилизинг-гормон лютеинизирующего гормона (РГЛГ) и его синтетические аналоги

Слайд 7Третьим уровнем регуляции- гипофиз
Гипофиз состоит из передней, задней и промежуточной
Третьим уровнем регуляции- гипофиз
Гипофиз состоит из передней, задней и промежуточной

Слайд 8Четвертому уровню регуляции- яичники
Функции яичника :
Генеративная функция (рост и созревание
Четвертому уровню регуляции- яичники
Функции яичника :
Генеративная функция (рост и созревание

Слайд 9Выделяют четыре типа фолликула:
Примордиальный фолликул состоит из незрелой яйцеклетки (ооцита) в профазе
Выделяют четыре типа фолликула:
Примордиальный фолликул состоит из незрелой яйцеклетки (ооцита) в профазе

Слайд 10В течение каждого менструального цикла от 3 до 30 примордиальных фолликулов начинают
В течение каждого менструального цикла от 3 до 30 примордиальных фолликулов начинают
Слайд 11Гормональная функция яичников
Рост, созревание фолликулов в яичниках и образование желтого тела сопровождаются
Гормональная функция яичников
Рост, созревание фолликулов в яичниках и образование желтого тела сопровождаются

Слайд 12Цикличность деятельности репродуктивной системы определяется принципами прямой и обратной связи, которая обеспечивается
Цикличность деятельности репродуктивной системы определяется принципами прямой и обратной связи, которая обеспечивается

Слайд 13Яичниковый цикл
Фолликулярная фаза:
Пульсирующая секреция и выделение ГнРГ приводят к высвобождению
Яичниковый цикл
Фолликулярная фаза:
Пульсирующая секреция и выделение ГнРГ приводят к высвобождению

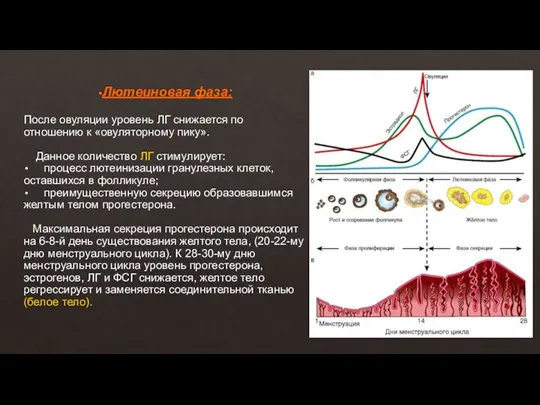
Слайд 14Лютеиновая фаза:
После овуляции уровень ЛГ снижается по отношению к «овуляторному пику».
Лютеиновая фаза:
После овуляции уровень ЛГ снижается по отношению к «овуляторному пику».
Слайд 15Пятый уровень регуляции органы-мишени: матка, маточные трубы, слизистая оболочка влагалища, а также
Пятый уровень регуляции органы-мишени: матка, маточные трубы, слизистая оболочка влагалища, а также

Слайд 16В эндометрии в течение менструального цикла происходят следующие изменения:
фаза десквамация и
В эндометрии в течение менструального цикла происходят следующие изменения:
фаза десквамация и

Слайд 17Фаза секреции (лютеиновая фаза в яичниках) связана с активностью желтого тела, длится
Фаза секреции (лютеиновая фаза в яичниках) связана с активностью желтого тела, длится

Слайд 18Менструация включает десквамацию, отторжение и регенерацию функционального слоя эндометрия.
В связи с
Менструация включает десквамацию, отторжение и регенерацию функционального слоя эндометрия.
В связи с

Слайд 19Регенерация эндометрия начинается сразу после отторжения некротизированного функционального слоя. Основой для регенерации
Регенерация эндометрия начинается сразу после отторжения некротизированного функционального слоя. Основой для регенерации

 Профилактика возникновения профессиональных заболеваний
Профилактика возникновения профессиональных заболеваний Проект психиатрической клиники Беды с башкой
Проект психиатрической клиники Беды с башкой Проект реабилитации детей-инвалидов посредством лечебной физкультуры Шанс
Проект реабилитации детей-инвалидов посредством лечебной физкультуры Шанс Препараты аминокислот, ферментов и антиферментов
Препараты аминокислот, ферментов и антиферментов Последствия аборта
Последствия аборта Аневризмы аорты. Определение
Аневризмы аорты. Определение Центральное, рефлекторное и местное, действие камфоры. Влияние на сердечно-сосудистую систему
Центральное, рефлекторное и местное, действие камфоры. Влияние на сердечно-сосудистую систему Несъёмные зубные протезы
Несъёмные зубные протезы Сифилис
Сифилис Влияние мобильных устройств на организм человека
Влияние мобильных устройств на организм человека Травма – основная причина смерти в популяции
Травма – основная причина смерти в популяции СВИЯЖ 30-150 рециркулятор бактерицидный
СВИЯЖ 30-150 рециркулятор бактерицидный Эпидемии в средние века и борьба с ними
Эпидемии в средние века и борьба с ними Иммунодефициты. Вторичные иммунодефициты
Иммунодефициты. Вторичные иммунодефициты Современная качественная колоноскопия в диагностике колоректального рака
Современная качественная колоноскопия в диагностике колоректального рака Первые признаки наркомании
Первые признаки наркомании скарлатина
скарлатина Инфекцияға қарсы (вирусқа және бактерияға қарсы) иммунитет механизмдері
Инфекцияға қарсы (вирусқа және бактерияға қарсы) иммунитет механизмдері Уход и наблюдение за детьми с заболеваниями ССС
Уход и наблюдение за детьми с заболеваниями ССС Почему важно соблюдать режим сна
Почему важно соблюдать режим сна Гибель клетки. Компенсаторно-приспособительные процессы. Регенерация, гипертрофия и гиперплазия
Гибель клетки. Компенсаторно-приспособительные процессы. Регенерация, гипертрофия и гиперплазия 14 Гуморальная и органы чувств 06d6354765eba1c9061e12e764083e0b
14 Гуморальная и органы чувств 06d6354765eba1c9061e12e764083e0b Пороки сердца
Пороки сердца Системная красная волчанка (СКВ)
Системная красная волчанка (СКВ) Интернист. Digest интерниста
Интернист. Digest интерниста Лечение повреждений вен. Методы восстановления
Лечение повреждений вен. Методы восстановления Применение метадоксила при токсическом поражении печени
Применение метадоксила при токсическом поражении печени Гоноре́я (от др.-греч. γόνος семенная жидкость и ῥέω теку)
Гоноре́я (от др.-греч. γόνος семенная жидкость и ῥέω теку)