- ПРОБЛЕМА ВЫДЕЛЕНИЯ CpG-ОСТРОВОВ И ГОМОЛОГИЧНЫХ ИМ СТРУКТУР В РАЗНЫХ ГРУППАХ ОРГАНИЗМОВ.

Содержание
- 2. CpG-islands in vertebrate genomes. M.Gardiner-Garden, M.Frommer (J.Mol.Biol.,1987, v.196. pp.261-282) Observed/Expected ratio > 0.60 Percent C +
- 3. Варианты расположения CpG-островов Связанные с промотором и первым экзоном Внутригенные, не захватывающие старт транскрипции Захватывающие последний
- 4. Assessment of clusters of transcription factor binding sites in relationship to human promoter, CpG islands and
- 5. Dualism of gene GC-content and CpG pattern in regards to expression in the human genome: magnitude
- 6. Наличие 5’-острова коррелирует с шириной спектра экспрессии и для растений. Gene-associated CpG Islands and the Expression
- 7. Comprehensive analysis of the base composition around the transcription start site in Metazoa Stein Aerts, Gert
- 8. Зачем нужны внутригенные острова? Genes and Transposons Are Differentially Methylated in Plants, but Not in Mammals
- 9. Distribution and Characterization of Regulatory Elements in the Human Genome (Jacek Majewski and Jurg Ott 2002,
- 11. Маевский и Отт – возможная роль CpG в сплайсинге, отсюда и падение в 3’-конце последнего экзона,
- 12. PEG3 (ZIM2) Импринтируемый ген. Обе изоформы – отцовские, но тканеспецифичность разная PEG3 - 7 первых экзонов
- 13. Determinants of CpG Islands: Expression in Early Embryo and Isochore Structure Loïc Ponger, Laurent Duret and
- 14. MEST- импринтированный ген Одна изоформа с отцовской экспрессией, другая – с биаллельной CpG-острова по всей видимости
- 15. CpG-острова в гене MEST (PEG1) у человека и мыши. У человека начало фрагмента в 2 т.п.н.
- 16. Выводы? Ужесточение критериев поиска не всегда целесообразно даже для 5’-концевых островов, хотя и позволяет отбросить повторы.
- 17. Структура CpG-островов и их флангов, связанных с альтернативными стартами, в гене MEST (PEG1) Скопления мотивов (локализовано
- 18. Биологическая роль продуктов локусов CDKN2a и CDKN2b
- 19. Гены CDKN2a и CDKN2b
- 20. р14 и р16 считываются с альтернативных стартов в разных рамках. CpG-острова, перекрывающиеся со стартом для р14,
- 21. Структура флангов CpG-острова, перекрывающегося с 1 α-экзоном гена CDKN2a Наиболее интересен «старый» остров, перекрывающийся со стартом
- 22. Структура CpG-островов локуса р15-р16 у человека, параметры поиска. комплемент Observed/Expected ratio > 0.60 Percent C +
- 23. Структура CpG-островов локуса р15-р16 у быка, параметры поиска. Комплемент Observed/Expected ratio > 0.60 Percent C +
- 24. Структура CpG-островов локуса р15-р16 у крысы, параметры поиска. Комплемент Observed/Expected ratio > 0.60 Percent C +
- 25. Тут опять ужесточение критериев поиска помешало бы увидеть биологически значимый, эволюционно более древний и консервативный CpG-остров,
- 26. Меланома у меченосцев связана с геном CDKNX, дупликация которого дала р15 и р16. По функциям ближе
- 27. Провоцируется межлинейными и межвидовыми скрещиваниями
- 28. Характеристики CpG-островов гена CDKNX у Xiphophorus helleri Вновь видим острова, перекрывающиеся со стартом р16 и с
- 29. Особенности CpG-островов у рыб Primordial stage? Острова короче, а повышения содержания G+C нет, хотя есть повышение
- 30. Возможна ли классификация островов по метилированности? CpG Island Methylation in Human Lymphocytes Is Highly Correlated with
- 31. Есть ли в других группах метилирование CpG и структуры, аналогичные CpG-островам? Если есть метилирование, то должно
- 32. Относительные частоты динуклеотида CpG у разных организмов. (Andrew J. Gentles and Samuel Karlin 2001 Genome Res
- 33. Метилирование у Drosophila melanogaster Есть высококонсервативная метилаза Dnmt2. Слабоактивна, метилирует в большей степени CpT и СрА,
- 34. Метилирование у других насекомых. Метилаза комара Anopheles gambiae очень похожа на таковую у дрозофилы. У молей
- 35. Comprehensive analysis of the base composition around the transcription start site in Metazoa У дрозофилы в
- 36. Особенности метилирования у растений. Есть не только ферменты, которые метилируют CpG, но также метилирующие СрNpG и
- 39. Для выделения CpG-островов у растений необходим пересчёт параметров для типичного для них %GC и отношения CpG/GpC.
- 40. По I.Ashikawa
- 42. Скачать презентацию
Слайд 2CpG-islands in vertebrate genomes.
M.Gardiner-Garden, M.Frommer
(J.Mol.Biol.,1987, v.196. pp.261-282)
Observed/Expected ratio > 0.60
Percent C
CpG-islands in vertebrate genomes.
M.Gardiner-Garden, M.Frommer
(J.Mol.Biol.,1987, v.196. pp.261-282)
Observed/Expected ratio > 0.60
Percent C

Слайд 3Варианты расположения CpG-островов
Связанные с промотором и первым экзоном
Внутригенные, не захватывающие старт транскрипции
Захватывающие
Варианты расположения CpG-островов
Связанные с промотором и первым экзоном
Внутригенные, не захватывающие старт транскрипции
Захватывающие

Слайд 4Assessment of clusters of transcription factor binding sites in
relationship to human promoter,
Assessment of clusters of transcription factor binding sites in relationship to human promoter,
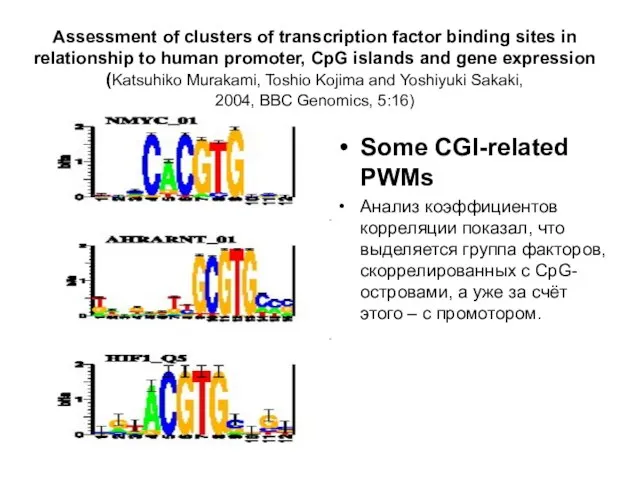
Слайд 5Dualism of gene GC-content and CpG pattern in regards to expression in
Dualism of gene GC-content and CpG pattern in regards to expression in

Слайд 6Наличие 5’-острова коррелирует с шириной спектра экспрессии и для растений.
Gene-associated CpG Islands
Наличие 5’-острова коррелирует с шириной спектра экспрессии и для растений.
Gene-associated CpG Islands

Слайд 7Comprehensive analysis of the base composition around the
transcription start site in Metazoa
Stein
Comprehensive analysis of the base composition around the transcription start site in Metazoa Stein
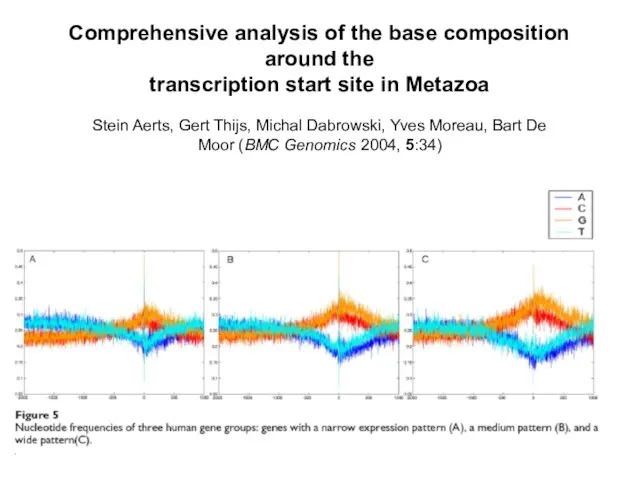
Слайд 8Зачем нужны внутригенные острова?
Genes and Transposons Are Differentially
Methylated in Plants, but Not
Зачем нужны внутригенные острова?
Genes and Transposons Are Differentially
Methylated in Plants, but Not

Слайд 9Distribution and Characterization of Regulatory
Elements in the Human Genome
(Jacek Majewski and Jurg
Distribution and Characterization of Regulatory Elements in the Human Genome (Jacek Majewski and Jurg
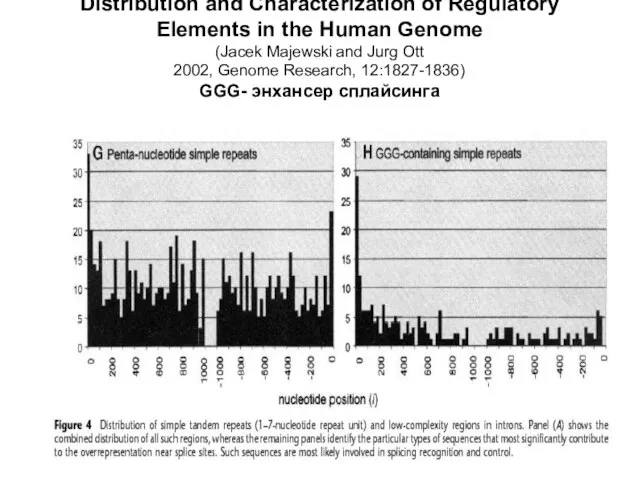
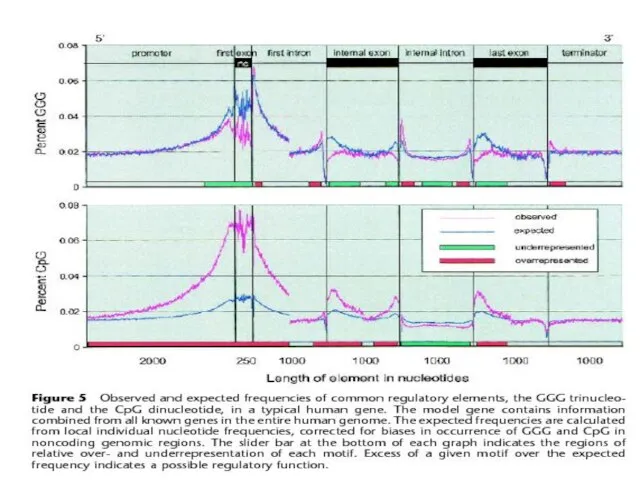
Слайд 11Маевский и Отт – возможная роль CpG в сплайсинге, отсюда и падение
Маевский и Отт – возможная роль CpG в сплайсинге, отсюда и падение

Слайд 12PEG3 (ZIM2)
Импринтируемый ген.
Обе изоформы – отцовские, но тканеспецифичность разная
PEG3 - 7
PEG3 (ZIM2)
Импринтируемый ген.
Обе изоформы – отцовские, но тканеспецифичность разная
PEG3 - 7

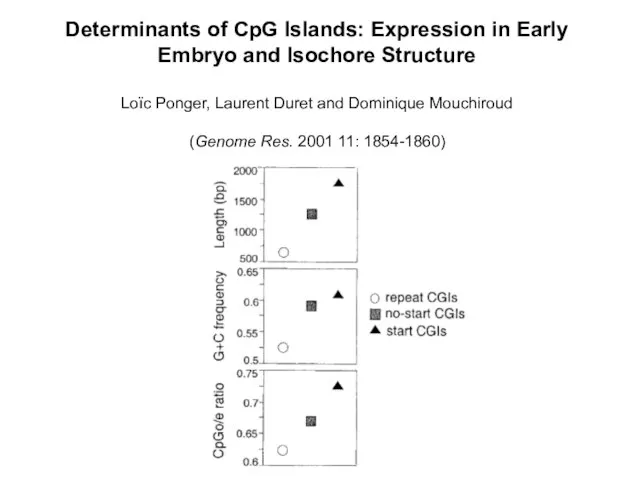
Слайд 13Determinants of CpG Islands: Expression in Early Embryo and Isochore Structure
Loïc Ponger,
Determinants of CpG Islands: Expression in Early Embryo and Isochore Structure Loïc Ponger,
Слайд 14MEST- импринтированный ген
Одна изоформа с отцовской экспрессией,
другая – с биаллельной
CpG-острова по
MEST- импринтированный ген
Одна изоформа с отцовской экспрессией,
другая – с биаллельной
CpG-острова по

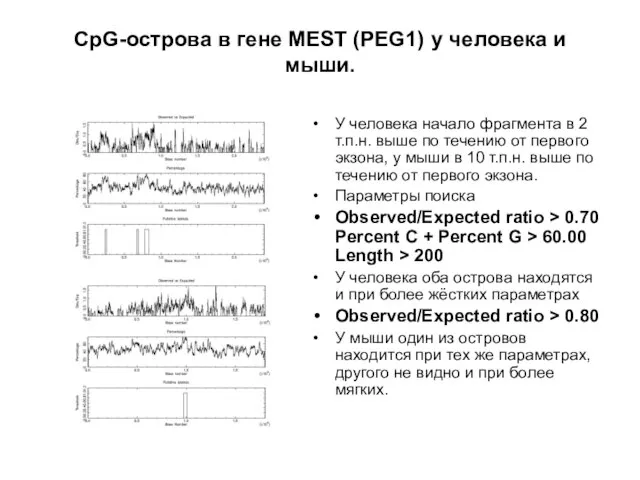
Слайд 15CpG-острова в гене MEST (PEG1) у человека и мыши.
У человека начало фрагмента
CpG-острова в гене MEST (PEG1) у человека и мыши.
У человека начало фрагмента
Слайд 16Выводы?
Ужесточение критериев поиска не всегда целесообразно даже для 5’-концевых островов, хотя и
Выводы?
Ужесточение критериев поиска не всегда целесообразно даже для 5’-концевых островов, хотя и

Слайд 17Структура CpG-островов и их флангов, связанных с альтернативными стартами,
в гене MEST
Структура CpG-островов и их флангов, связанных с альтернативными стартами, в гене MEST

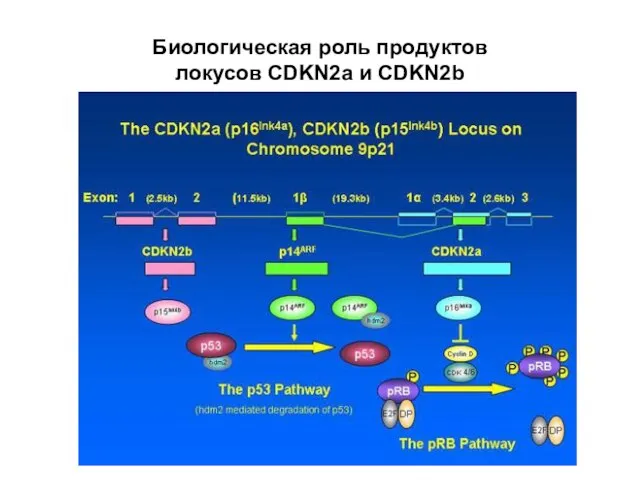
Слайд 18Биологическая роль продуктов
локусов CDKN2a и CDKN2b
Биологическая роль продуктов
локусов CDKN2a и CDKN2b
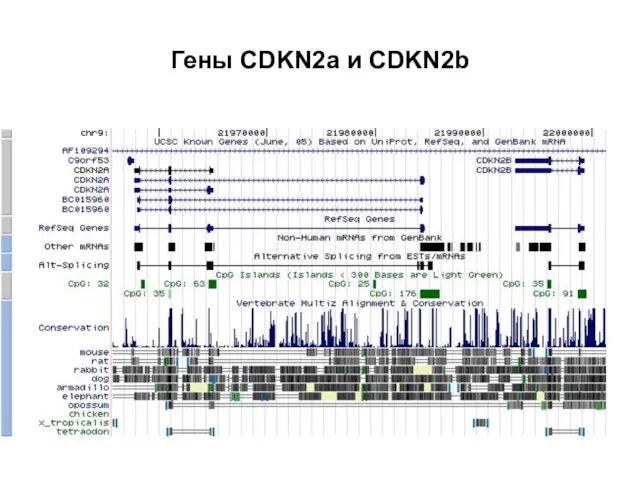
Слайд 19Гены CDKN2a и CDKN2b
Гены CDKN2a и CDKN2b
Слайд 20р14 и р16 считываются с альтернативных стартов в разных рамках. CpG-острова, перекрывающиеся
р14 и р16 считываются с альтернативных стартов в разных рамках. CpG-острова, перекрывающиеся

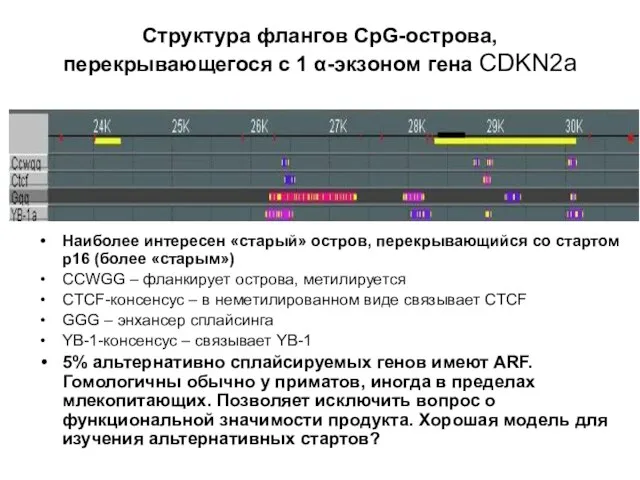
Слайд 21Структура флангов CpG-острова, перекрывающегося с 1 α-экзоном гена CDKN2a
Наиболее интересен «старый» остров,
Структура флангов CpG-острова, перекрывающегося с 1 α-экзоном гена CDKN2a
Наиболее интересен «старый» остров,
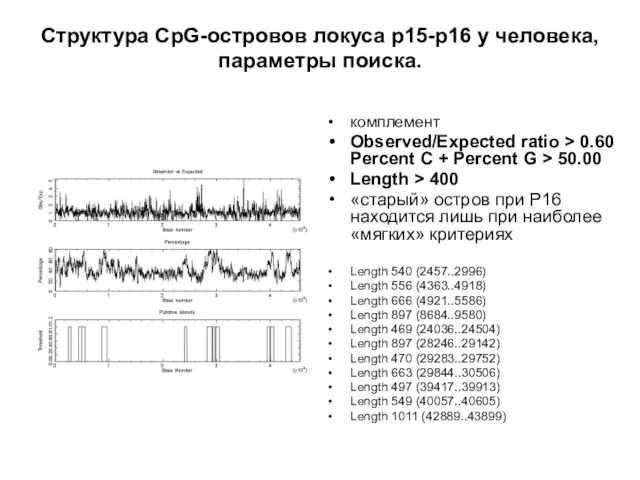
Слайд 22Структура CpG-островов локуса р15-р16 у человека,
параметры поиска.
комплемент
Observed/Expected ratio > 0.60 Percent
Структура CpG-островов локуса р15-р16 у человека,
параметры поиска.
комплемент
Observed/Expected ratio > 0.60 Percent
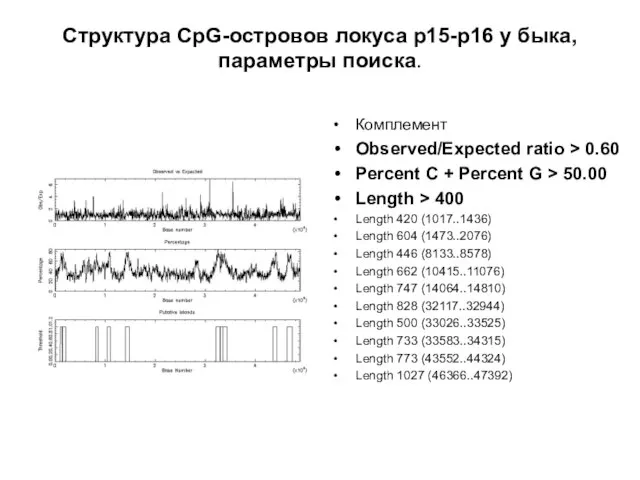
Слайд 23Структура CpG-островов локуса р15-р16 у быка,
параметры поиска.
Комплемент
Observed/Expected ratio > 0.60
Percent
Структура CpG-островов локуса р15-р16 у быка,
параметры поиска.
Комплемент
Observed/Expected ratio > 0.60
Percent
Слайд 24Структура CpG-островов локуса р15-р16 у крысы,
параметры поиска.
Комплемент
Observed/Expected ratio > 0.60 Percent
Структура CpG-островов локуса р15-р16 у крысы,
параметры поиска.
Комплемент
Observed/Expected ratio > 0.60 Percent

Слайд 25Тут опять ужесточение критериев поиска помешало бы увидеть биологически значимый, эволюционно более
Тут опять ужесточение критериев поиска помешало бы увидеть биологически значимый, эволюционно более

Слайд 26Меланома у меченосцев
связана с геном CDKNX, дупликация которого дала р15 и р16.
Меланома у меченосцев связана с геном CDKNX, дупликация которого дала р15 и р16.

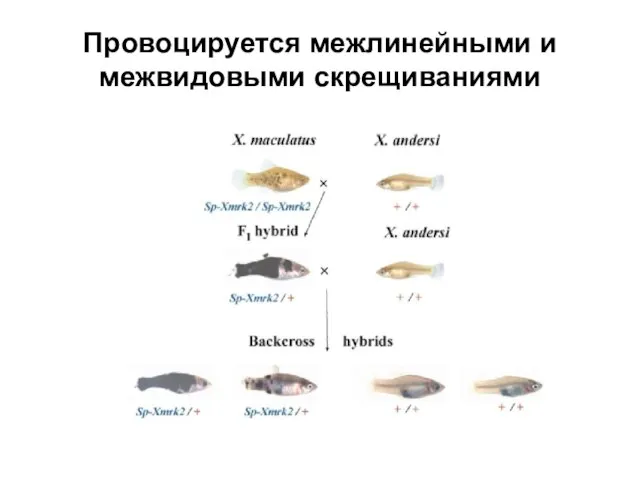
Слайд 27Провоцируется межлинейными и межвидовыми скрещиваниями
Провоцируется межлинейными и межвидовыми скрещиваниями
Слайд 28Характеристики CpG-островов гена CDKNX у Xiphophorus helleri
Вновь видим острова, перекрывающиеся со стартом
Характеристики CpG-островов гена CDKNX у Xiphophorus helleri Вновь видим острова, перекрывающиеся со стартом

Слайд 29Особенности CpG-островов у рыб
Primordial stage?
Острова короче, а повышения содержания G+C нет,
Особенности CpG-островов у рыб
Primordial stage?
Острова короче, а повышения содержания G+C нет,

Слайд 30
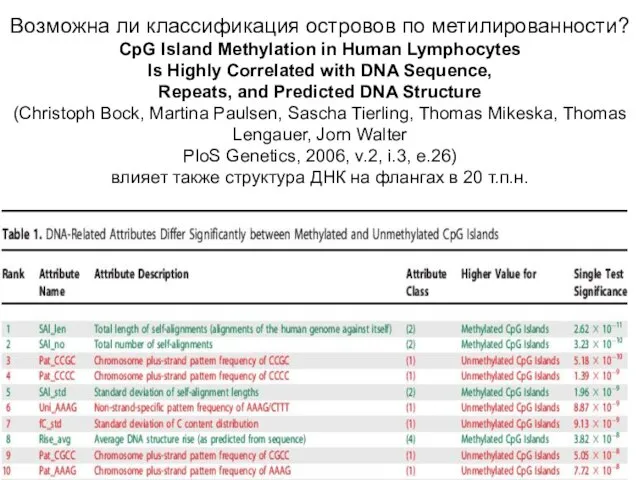
Возможна ли классификация островов по метилированности?
CpG Island Methylation in Human Lymphocytes
Is Highly
Возможна ли классификация островов по метилированности? CpG Island Methylation in Human Lymphocytes Is Highly
Слайд 31Есть ли в других группах метилирование CpG и структуры, аналогичные CpG-островам?
Если есть
Есть ли в других группах метилирование CpG и структуры, аналогичные CpG-островам? Если есть

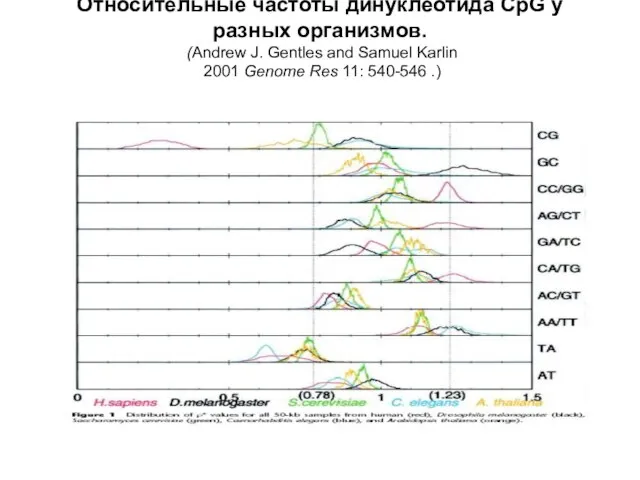
Слайд 32Относительные частоты динуклеотида CpG у разных организмов.
(Andrew J. Gentles and Samuel
Относительные частоты динуклеотида CpG у разных организмов. (Andrew J. Gentles and Samuel
Слайд 33Метилирование у
Drosophila melanogaster
Есть высококонсервативная метилаза Dnmt2. Слабоактивна, метилирует в большей степени
Метилирование у
Drosophila melanogaster
Есть высококонсервативная метилаза Dnmt2. Слабоактивна, метилирует в большей степени

Слайд 34Метилирование у других насекомых.
Метилаза комара Anopheles gambiae очень похожа на таковую у
Метилирование у других насекомых.
Метилаза комара Anopheles gambiae очень похожа на таковую у

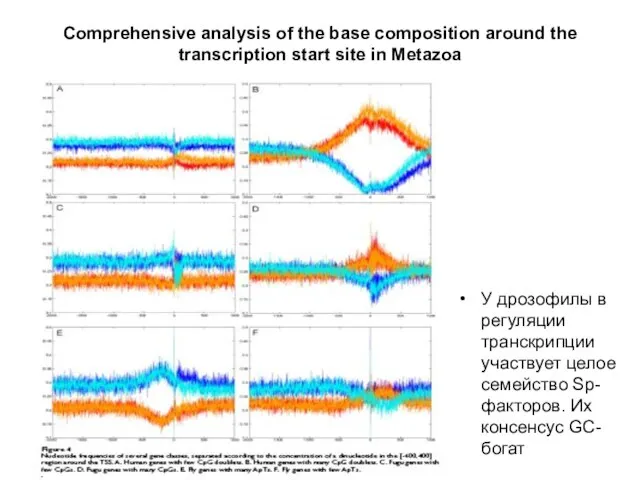
Слайд 35Comprehensive analysis of the base composition around the
transcription start site in Metazoa
У
Comprehensive analysis of the base composition around the
transcription start site in Metazoa
У
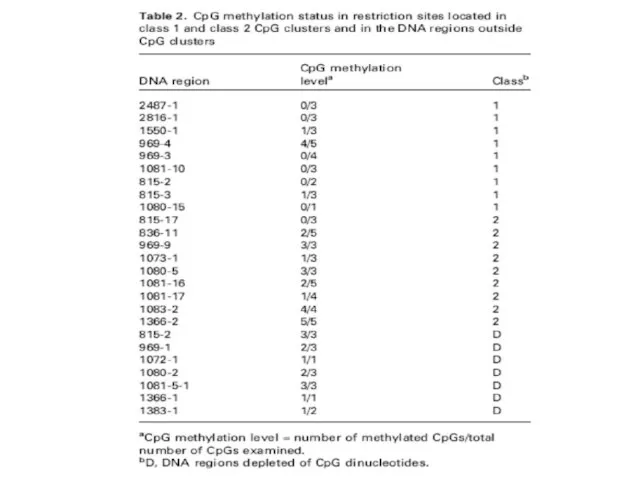
Слайд 36Особенности метилирования у растений.
Есть не только ферменты, которые метилируют CpG, но также
Особенности метилирования у растений.
Есть не только ферменты, которые метилируют CpG, но также


Слайд 39Для выделения CpG-островов у растений необходим пересчёт параметров для типичного для них
Для выделения CpG-островов у растений необходим пересчёт параметров для типичного для них

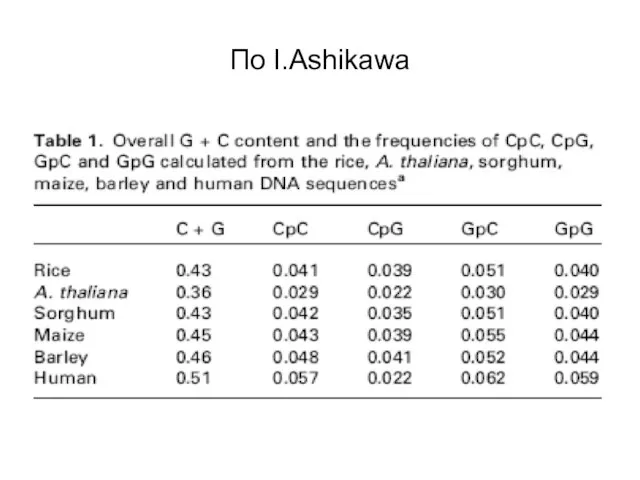
Слайд 40По I.Ashikawa
По I.Ashikawa
 Психика человека
Психика человека ВОССТАНОВИТЕЛЬНОЕ ПРАВОСУДИЕ иЮВЕНАЛЬНЫЕ ТЕХНОЛОГИИ в РОССИИ
ВОССТАНОВИТЕЛЬНОЕ ПРАВОСУДИЕ иЮВЕНАЛЬНЫЕ ТЕХНОЛОГИИ в РОССИИ МЕЖВЕДОМСТВЕННОЕ ВЗАИМОДЕЙСТВИЕ ПРИ ОРГАНИЗАЦИИ СЕМЕЙНОГО УСТРОЙСТВА ДЕТЕЙ,ОСТАВШИХСЯБЕЗ ПОПЕЧЕНИЯ РОДИТЕЛЕЙ
МЕЖВЕДОМСТВЕННОЕ ВЗАИМОДЕЙСТВИЕ ПРИ ОРГАНИЗАЦИИ СЕМЕЙНОГО УСТРОЙСТВА ДЕТЕЙ,ОСТАВШИХСЯБЕЗ ПОПЕЧЕНИЯ РОДИТЕЛЕЙ Назначение и структура бизнес-плана
Назначение и структура бизнес-плана Тонкости продвижения интернет-магазиновв поисковых системах
Тонкости продвижения интернет-магазиновв поисковых системах Євроінтеграція України як чинник соціально-економічного розвитку держави. Роль освіти в розвитку партнерства України з іншими де
Євроінтеграція України як чинник соціально-економічного розвитку держави. Роль освіти в розвитку партнерства України з іншими де Особенности модернизации России – процесс взаимодействия инновационного и сырьевого векторов экономики Карпова Анна Владимиров
Особенности модернизации России – процесс взаимодействия инновационного и сырьевого векторов экономики Карпова Анна Владимиров Развитие психики человека
Развитие психики человека Сатиры А.Д.Кантемира
Сатиры А.Д.Кантемира Конкурсный проект смотровой площадки на вершине горы Машук
Конкурсный проект смотровой площадки на вершине горы Машук Вводный инструктаж. Формирование команды
Вводный инструктаж. Формирование команды Новая экономичная система импульсного пневмотранспорта порошкообразных сред
Новая экономичная система импульсного пневмотранспорта порошкообразных сред Подбор конфигурации и модернизация средств вычислительной техники
Подбор конфигурации и модернизация средств вычислительной техники Школьный Художественный музей
Школьный Художественный музей Прикладная геоэкология
Прикладная геоэкология Подвижная игрушка Слоненок
Подвижная игрушка Слоненок Я - мэр города Петрозаводска
Я - мэр города Петрозаводска Презентация на тему Иисус Христос – историческая личность или мифологический герой
Презентация на тему Иисус Христос – историческая личность или мифологический герой Культура Древнего Китая
Культура Древнего Китая Отраслевое административно-правовое регулирование в хозяйственно-экономических комплексах [часть 2]
Отраслевое административно-правовое регулирование в хозяйственно-экономических комплексах [часть 2] Свойства жидкостей, газов и твердых тел в пословицах
Свойства жидкостей, газов и твердых тел в пословицах Презентация на тему Богомол
Презентация на тему Богомол Временное трудоустройство. Подростки
Временное трудоустройство. Подростки Свобода. Уверенность. Выгода.
Свобода. Уверенность. Выгода. Комаров Сергей Приложения для социальных сетей. Использование приложений в качестве рекламных инструментов.
Комаров Сергей Приложения для социальных сетей. Использование приложений в качестве рекламных инструментов. Презентация "Религиозные праздники христиан" - скачать презентации по МХК
Презентация "Религиозные праздники христиан" - скачать презентации по МХК Оценка необходимости и обоснованности внедрения системы CRM в компании АБВ
Оценка необходимости и обоснованности внедрения системы CRM в компании АБВ Искусство быть здоровым.
Искусство быть здоровым.