- Регуляция работы генов

Содержание
- 2. Классификация генов Структурные гены несут информацию о белках-ферментах и гистонах, о последовательности нуклеотидов в различных видах
- 3. Генотип у всех соматических клеток одинаковый (следствие равного распределения генетического материала между дочерними клетками при митозе),
- 4. Регуляция работы генов Некоторые ферменты у дрожжей и бактерий образуются в клетках только при выращивании их
- 5. Регуляция работы генов у прокариот Схема регуляции транскрипции у прокариот была предложена Ф. Жакобом и Ж.
- 6. Регуляция работы генов у прокариот Если в клетку поступает индуктор, то он связывает белок-репрессор (вступает с
- 7. Регуляция работы генов у эукариот Схема регуляции транскрипции у эукариот разработана Г. П. Георгиевым (1972). Принцип
- 8. Регуляция работы генов у эукариот Работу транскриптона регулирует несколько генов-регуляторов, дающих информацию для синтеза нескольких белков-репрессоров.
- 9. Регуляция работы генов у эукариот В дальнейшем в ядре клетки происходит процессинг - ферментативное разрушение неинформативной
- 10. Регуляция работы генов у эукариот В геноме эукариот встречаются уникальные последовательности нуклеотидов (одна в геноме), составляющие
- 11. Механизмы реализации генетической информации Чтобы понять механизмы реализации генетической информации, необходимо дополнить центральную догму молекулярной биологии.
- 12. В настоящее время центральная догма молекулярной биологии может быть представлена следующей схемой Репликация ДНК и-РНК белок
- 13. Механизмы реализации генетической информации Механизм дифференцировки стволовых клеток можно представить следующим образом. Недифференцированные клетки имеют разный
- 14. Механизмы реализации генетической информации Условно гены можно разделить на три группы: функционирующие во всех клетках (например,
- 15. Цитоплазматическая наследственность Наряду с ядерными генами, локализованными в хромосомах, обнаружены факторы наследственности, находящиеся в цитоплазме. Их
- 16. Цитоплазматическая наследственность Возможными критериями цитоплазматической наследственности являются: отсутствие количественного менделевского расщепления в потомстве; невозможность выявления сцепления;
- 17. Цитоплазматическая наследственность Открытие пластидной наследственности принадлежит К. Корренсу (1908), описавшему пестролистность у растения "ночная красавица". У
- 18. Цитоплазматическая наследственность Митохондриальная наследственность описана Б. Эфрусси (1949). Около 1% хлебных дрожжей дают карликовые колонии. Оказалось,
- 19. Цитоплазматическая наследственность Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16 569 пар нуклеотидов. В состав
- 21. Скачать презентацию
Слайд 2Классификация генов
Структурные гены несут информацию о белках-ферментах и гистонах, о последовательности
Классификация генов
Структурные гены несут информацию о белках-ферментах и гистонах, о последовательности

Слайд 3 Генотип у всех соматических клеток одинаковый
(следствие равного распределения генетического
материала
Генотип у всех соматических клеток одинаковый
(следствие равного распределения генетического
материала

Слайд 4Регуляция работы генов
Некоторые ферменты у дрожжей и бактерий образуются в клетках
Регуляция работы генов
Некоторые ферменты у дрожжей и бактерий образуются в клетках

Слайд 5Регуляция работы генов у прокариот
Схема регуляции транскрипции у прокариот была предложена
Регуляция работы генов у прокариот
Схема регуляции транскрипции у прокариот была предложена

Слайд 6Регуляция работы генов у прокариот
Если в клетку поступает индуктор, то он связывает
Регуляция работы генов у прокариот
Если в клетку поступает индуктор, то он связывает

Слайд 7Регуляция работы генов у эукариот
Схема регуляции транскрипции у эукариот разработана Г.
Регуляция работы генов у эукариот
Схема регуляции транскрипции у эукариот разработана Г.

Слайд 8Регуляция работы генов у эукариот
Работу транскриптона регулирует несколько генов-регуляторов, дающих информацию для
Регуляция работы генов у эукариот
Работу транскриптона регулирует несколько генов-регуляторов, дающих информацию для

Слайд 9Регуляция работы генов у эукариот
В дальнейшем в ядре клетки происходит процессинг -
Регуляция работы генов у эукариот
В дальнейшем в ядре клетки происходит процессинг -

Слайд 10Регуляция работы генов у эукариот
В геноме эукариот встречаются уникальные последовательности нуклеотидов (одна
Регуляция работы генов у эукариот
В геноме эукариот встречаются уникальные последовательности нуклеотидов (одна

Слайд 11Механизмы реализации генетической информации
Чтобы понять механизмы реализации генетической информации, необходимо дополнить
Механизмы реализации генетической информации
Чтобы понять механизмы реализации генетической информации, необходимо дополнить

Слайд 12В настоящее время центральная догма молекулярной биологии может быть представлена следующей схемой
Репликация
В настоящее время центральная догма молекулярной биологии может быть представлена следующей схемой
Репликация
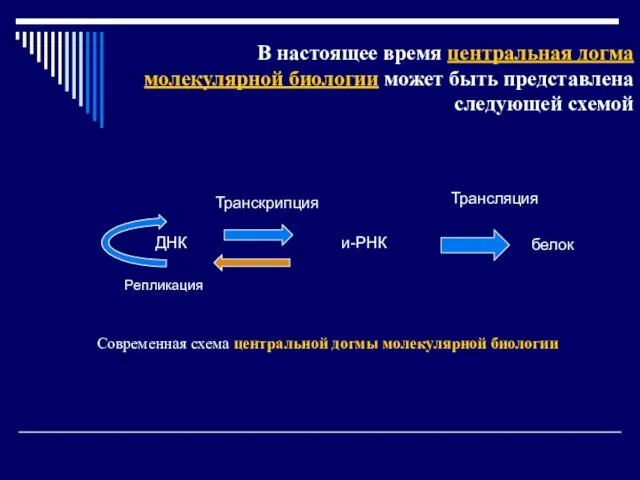
Слайд 13Механизмы реализации генетической информации
Механизм дифференцировки стволовых клеток можно представить следующим образом. Недифференцированные
Механизмы реализации генетической информации
Механизм дифференцировки стволовых клеток можно представить следующим образом. Недифференцированные

Слайд 14Механизмы реализации генетической информации
Условно гены можно разделить на три группы:
функционирующие во всех
Механизмы реализации генетической информации
Условно гены можно разделить на три группы:
функционирующие во всех

Слайд 15Цитоплазматическая наследственность
Наряду с ядерными генами, локализованными в хромосомах, обнаружены факторы наследственности,
Цитоплазматическая наследственность
Наряду с ядерными генами, локализованными в хромосомах, обнаружены факторы наследственности,

Слайд 16Цитоплазматическая наследственность
Возможными критериями цитоплазматической наследственности являются:
отсутствие количественного менделевского расщепления в потомстве;
невозможность выявления
Цитоплазматическая наследственность
Возможными критериями цитоплазматической наследственности являются:
отсутствие количественного менделевского расщепления в потомстве;
невозможность выявления

Слайд 17Цитоплазматическая наследственность
Открытие пластидной наследственности принадлежит К. Корренсу (1908), описавшему пестролистность у растения
Цитоплазматическая наследственность
Открытие пластидной наследственности принадлежит К. Корренсу (1908), описавшему пестролистность у растения

Слайд 18Цитоплазматическая наследственность
Митохондриальная наследственность описана Б. Эфрусси (1949).
Около 1% хлебных дрожжей дают
Цитоплазматическая наследственность
Митохондриальная наследственность описана Б. Эфрусси (1949).
Около 1% хлебных дрожжей дают

Слайд 19Цитоплазматическая наследственность
Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16 569 пар
Цитоплазматическая наследственность
Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16 569 пар

 Тип Кишечнополостные животные
Тип Кишечнополостные животные Сервировка стола к завтраку
Сервировка стола к завтраку Архитектурные постройки
Архитектурные постройки Макияж при близорукости, дальнозоркости, для тех, кто носит линзы
Макияж при близорукости, дальнозоркости, для тех, кто носит линзы الخطأ السياقي
الخطأ السياقي Презентация по обществознанию на тему _Мировоззрение и его роль в жизни человека_ (10
Презентация по обществознанию на тему _Мировоззрение и его роль в жизни человека_ (10 Spotlight
Spotlight Создай образ с TRESemme
Создай образ с TRESemme Linkin park
Linkin park Презентация на тему Степные животные
Презентация на тему Степные животные Сочинение –описание животного.Моя собака Крош.
Сочинение –описание животного.Моя собака Крош. Предпринимательский модуль. Мастерская агробизнеса
Предпринимательский модуль. Мастерская агробизнеса Обзор методологии Scrum Auriga Inc. Дмитрий Сидоренко. - презентация
Обзор методологии Scrum Auriga Inc. Дмитрий Сидоренко. - презентация Обращение к психологу, страшно или нет
Обращение к психологу, страшно или нет Салон штор и гардин LINENS™
Салон штор и гардин LINENS™ Непроектное мероприятие «Образование детей-инвалидов»
Непроектное мероприятие «Образование детей-инвалидов» Гимнастика. Виды гимнастики
Гимнастика. Виды гимнастики Балет - один из языков истории
Балет - один из языков истории Отзывы о Чувышии
Отзывы о Чувышии Сигналы тревоги.
Сигналы тревоги. Олимпиаде в Сочи посвящается… Кроссворд Олимпийские игры
Олимпиаде в Сочи посвящается… Кроссворд Олимпийские игры Современный дизайн. Место дизайна в современном мире. Тренды в графическом дизайне
Современный дизайн. Место дизайна в современном мире. Тренды в графическом дизайне Время Present Perfect
Время Present Perfect Бережливое управление МБУ УБГБ (1)
Бережливое управление МБУ УБГБ (1) Лавис и резерваж. Манеры офорта
Лавис и резерваж. Манеры офорта Презентация на тему Фотосинтез и дыхание растений (6 класс)
Презентация на тему Фотосинтез и дыхание растений (6 класс)  Страна под настроение
Страна под настроение Создание системы бюджетного управления
Создание системы бюджетного управления