- Биохимические особенности течения злокачественных процессов

Содержание
- 2. АКТУАЛЬНОСТЬ Актуальность возможности использования показателей протеолитических систем для оценки прогноза продиктована доказательством их роли на этапах
- 3. АКТУАЛЬНОСТЬ Поэтому проблема персонифицированного подхода к лечению онкологических больных имеет большое научное и практическое значение. Поиск
- 4. РЕЗУЛЬТАТЫ Фенотип злокачественной опухоли формируется в результате изменения содержания патогенетически значимых белков в клетках, на которое
- 5. РЕЗУЛЬТАТЫ На биологические характеристики опухоли может повлиять постоянное изменение протеома, которое происходит под влиянием различных факторов
- 6. РЕЗУЛЬТАТЫ Одним из основных путей регуляции состава и качества протеома является протеолиз, опосредуемый убиквитин-протеасомной системой. Протеасомная
- 7. РЕЗУЛЬТАТЫ Каталитические комплексы протеасомной системы представлены двумя пулами: 20S- и 26S-протеаcомы. Непосредственно протеолиз происходит в каталитическом
- 8. РЕЗУЛЬТАТЫ Связывание регуляторной частицы (11S, 19S, РА200) с а-кольцами контролирует состояние ворот и способствует многократному увеличению
- 9. РЕЗУЛЬТАТЫ Протеасомы играют важную роль в патогенезе злокачественных опухолей. Продвижение клетки по клеточному циклу регулируется специфическими
- 10. РЕЗУЛЬТАТЫ Убиквитин-протеасомная система принимает участие как в разрушении самих циклинов, их комплексов, так и в регуляции
- 11. РЕЗУЛЬТАТЫ Многие ядерные белки, опосредующие программируемую клеточную гибель, являются субстратами для протеасом: транскрипционные факторы (c-Myc, AP-1),
- 12. РЕЗУЛЬТАТЫ Другим важным молекулярным механизмом развития и прогрессирования рака является внутриклеточный протеолиз, опосредованный кальпаиновой системой. Кальпаины
- 13. РЕЗУЛЬТАТЫ Кальпаиновая система деградации белков также осуществляет разрушение большого количества внутриклеточных белков, в том числе различных
- 15. Скачать презентацию
Слайд 2АКТУАЛЬНОСТЬ
Актуальность возможности использования показателей протеолитических систем для оценки прогноза продиктована доказательством их
АКТУАЛЬНОСТЬ
Актуальность возможности использования показателей протеолитических систем для оценки прогноза продиктована доказательством их
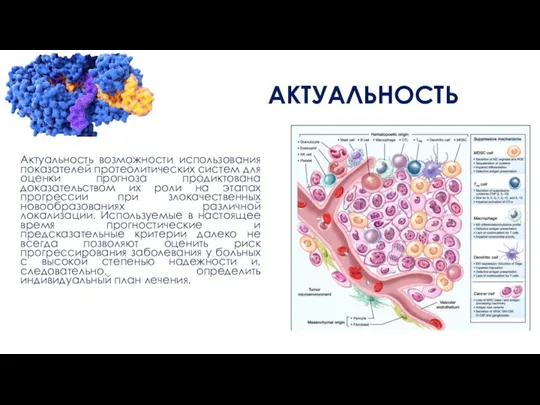
Слайд 3АКТУАЛЬНОСТЬ
Поэтому проблема персонифицированного подхода к лечению онкологических больных имеет большое научное и
АКТУАЛЬНОСТЬ
Поэтому проблема персонифицированного подхода к лечению онкологических больных имеет большое научное и

Слайд 4РЕЗУЛЬТАТЫ
Фенотип злокачественной опухоли формируется в результате изменения содержания патогенетически значимых белков в
РЕЗУЛЬТАТЫ
Фенотип злокачественной опухоли формируется в результате изменения содержания патогенетически значимых белков в
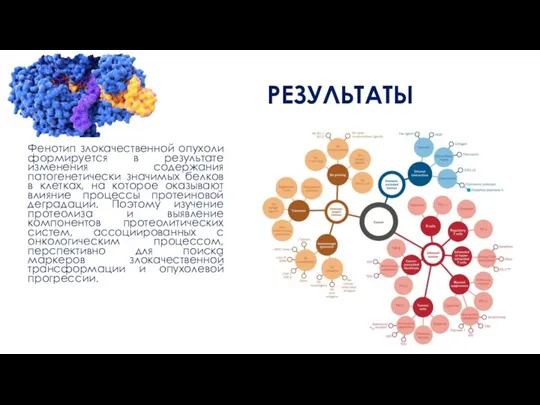
Слайд 5РЕЗУЛЬТАТЫ
На биологические характеристики опухоли может повлиять постоянное изменение протеома, которое происходит под
РЕЗУЛЬТАТЫ
На биологические характеристики опухоли может повлиять постоянное изменение протеома, которое происходит под
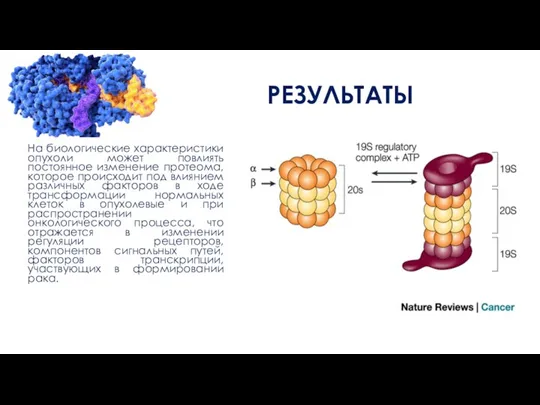
Слайд 6РЕЗУЛЬТАТЫ
Одним из основных путей регуляции состава и качества протеома является протеолиз, опосредуемый
РЕЗУЛЬТАТЫ
Одним из основных путей регуляции состава и качества протеома является протеолиз, опосредуемый
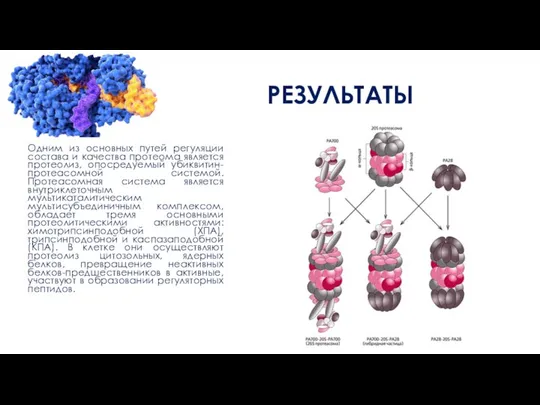
Слайд 7РЕЗУЛЬТАТЫ
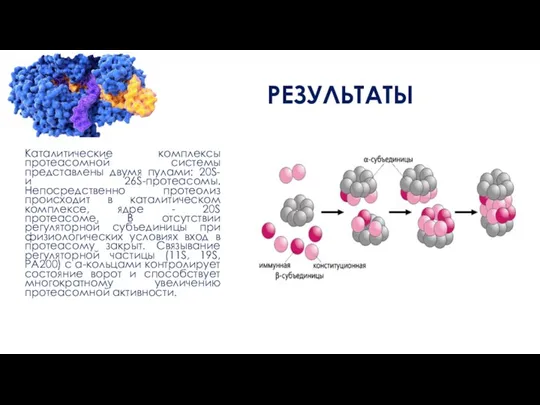
Каталитические комплексы протеасомной системы представлены двумя пулами: 20S- и 26S-протеаcомы. Непосредственно протеолиз
РЕЗУЛЬТАТЫ
Каталитические комплексы протеасомной системы представлены двумя пулами: 20S- и 26S-протеаcомы. Непосредственно протеолиз
Слайд 8РЕЗУЛЬТАТЫ
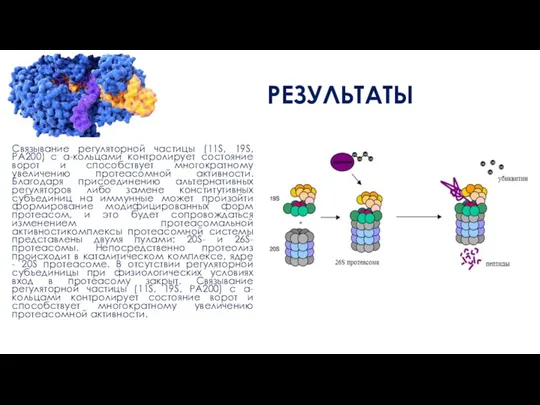
Связывание регуляторной частицы (11S, 19S, РА200) с а-кольцами контролирует состояние ворот и
РЕЗУЛЬТАТЫ
Связывание регуляторной частицы (11S, 19S, РА200) с а-кольцами контролирует состояние ворот и
Слайд 9РЕЗУЛЬТАТЫ
Протеасомы играют важную роль в патогенезе злокачественных опухолей. Продвижение клетки по клеточному
РЕЗУЛЬТАТЫ
Протеасомы играют важную роль в патогенезе злокачественных опухолей. Продвижение клетки по клеточному

Слайд 10РЕЗУЛЬТАТЫ
Убиквитин-протеасомная система принимает участие как в разрушении самих циклинов, их комплексов, так
РЕЗУЛЬТАТЫ
Убиквитин-протеасомная система принимает участие как в разрушении самих циклинов, их комплексов, так

Слайд 11РЕЗУЛЬТАТЫ
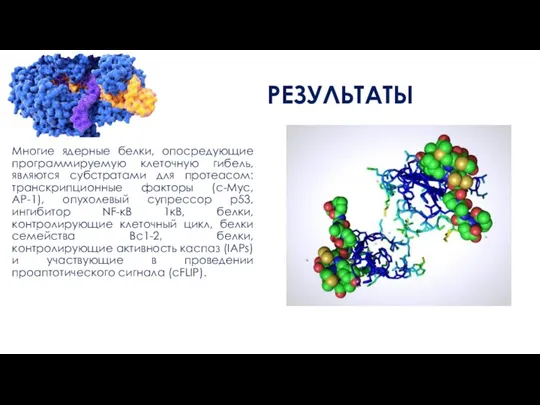
Многие ядерные белки, опосредующие программируемую клеточную гибель, являются субстратами для протеасом: транскрипционные
РЕЗУЛЬТАТЫ
Многие ядерные белки, опосредующие программируемую клеточную гибель, являются субстратами для протеасом: транскрипционные
Слайд 12РЕЗУЛЬТАТЫ
Другим важным молекулярным механизмом развития и прогрессирования рака является внутриклеточный протеолиз, опосредованный
РЕЗУЛЬТАТЫ
Другим важным молекулярным механизмом развития и прогрессирования рака является внутриклеточный протеолиз, опосредованный

Слайд 13РЕЗУЛЬТАТЫ
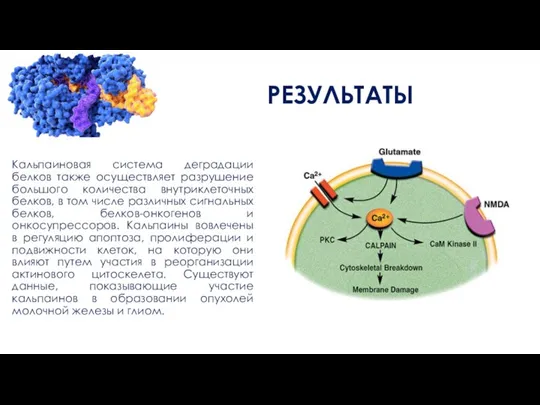
Кальпаиновая система деградации белков также осуществляет разрушение большого количества внутриклеточных белков, в
РЕЗУЛЬТАТЫ
Кальпаиновая система деградации белков также осуществляет разрушение большого количества внутриклеточных белков, в
 Дифтерия
Дифтерия Микробиологические трансформации органических соединений
Микробиологические трансформации органических соединений Заболевания, передающиеся половым путем и меры их профилактики
Заболевания, передающиеся половым путем и меры их профилактики Введение. Развитие фармации Свердловской области. Лекция №1
Введение. Развитие фармации Свердловской области. Лекция №1 Химиотерапевтические препараты. Антибиотики. Противовирусные средства
Химиотерапевтические препараты. Антибиотики. Противовирусные средства Этические вопросы реанимации новорожденных детей
Этические вопросы реанимации новорожденных детей Гуморальная регуляция
Гуморальная регуляция Фетоплацентарная недостаточность. Этиология, классификация, клиника
Фетоплацентарная недостаточность. Этиология, классификация, клиника Межклеточные взаимодействия через белки суперсемейства иммуноглобулинов
Межклеточные взаимодействия через белки суперсемейства иммуноглобулинов Лучевая диагностика опорно - двигательной системы. Повреждения костей и суставов (перелом, вывих)
Лучевая диагностика опорно - двигательной системы. Повреждения костей и суставов (перелом, вывих) Членовредительство. Самоповреждение
Членовредительство. Самоповреждение Наследственные болезни. МГК. /практический навык/
Наследственные болезни. МГК. /практический навык/ Дигитальная субтракционная ангиография
Дигитальная субтракционная ангиография Качество медико-социальных услуг, оказываемых пожилому населению
Качество медико-социальных услуг, оказываемых пожилому населению Прикладная физика кровообращения
Прикладная физика кровообращения Диспансеризция детей от 1 года до 3 лет
Диспансеризция детей от 1 года до 3 лет Қазіргі таңдағы оқу-тәрбие үдерісіндегі тәрбие технологиясы
Қазіргі таңдағы оқу-тәрбие үдерісіндегі тәрбие технологиясы Болезни органов дыхания
Болезни органов дыхания Особенности лекарственной терапии в гериатрии. Лекция 5
Особенности лекарственной терапии в гериатрии. Лекция 5 Программа физической реабилитации пациентов 5-6 лет с врожденной спинномозговой грыжей менингоцеле на стационарном этапе
Программа физической реабилитации пациентов 5-6 лет с врожденной спинномозговой грыжей менингоцеле на стационарном этапе ВИЧ-инфекция
ВИЧ-инфекция Здоровое питание - активное долголетие
Здоровое питание - активное долголетие Знаки медицинского и санитарного назначения
Знаки медицинского и санитарного назначения Кардиоренальный анемический синдром
Кардиоренальный анемический синдром Железодефицитные синдромы у больных с колоректальным раком до лечения
Железодефицитные синдромы у больных с колоректальным раком до лечения Принципы организации работы ПЦР-лаборатории
Принципы организации работы ПЦР-лаборатории Лептоспироз. Збудник
Лептоспироз. Збудник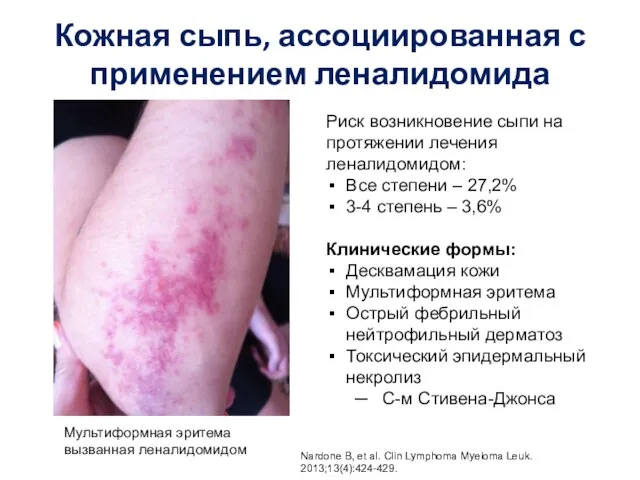 Кожная сыпь
Кожная сыпь