- Лекция_10

Содержание
- 2. Иммунодефициты Первичные Вторичные (врожденные) (приобретенные) Неспособность высшего организма противостоять инфекции Избегание патогеном действия иммунной системы
- 3. Многие инфекционные агенты существуют в нескольких антигенных вариантах. Streptococcus pneumoniae – 84 серотипа = 84 варианта
- 4. Множественные антигенные варианты полисахаридной капсулы Streptococcus pneumoniae позволяют бактерии вызывать повторную инфекцию у уже переболевшего индивидуума
- 5. Антигенный дрейф (antigenic drift) и антигенный шифт/сдвиг (antigenic shift) у вируса гриппа Antigenic drift антигенный дрейф
- 6. Для вируса гриппа типа А
- 7. Смена подтипов вируса гриппа А и связанные с ней пандемии и эпидемии человека Антигенный шифт вызывает
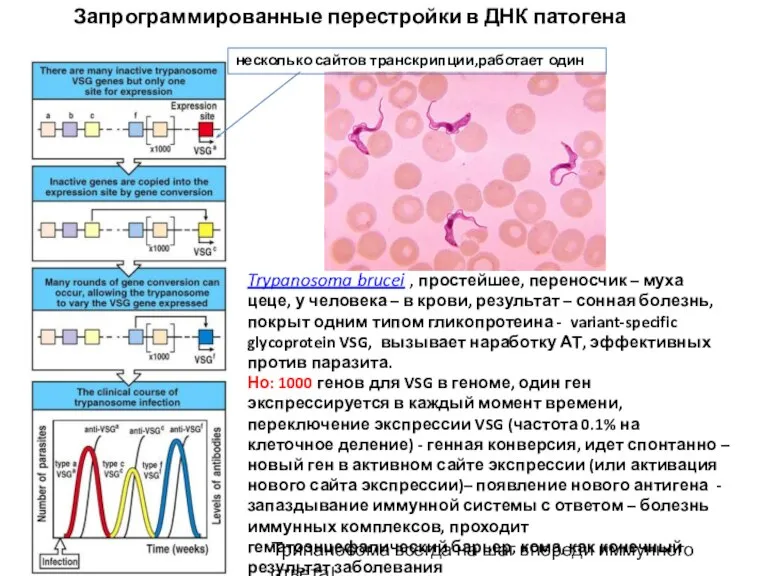
- 8. Запрограммированные перестройки в ДНК патогена Trypanosoma brucei , простейшее, переносчик – муха цеце, у человека –
- 9. Бактерии имеют адгезины, позволяющие им или просто присоединяться к клетке (Neisseria gonorrhoeae, белок адгезии - пилин)
- 10. Некоторые вирусы могут «исчезать» из видимости иммунной системы в латентный период - персистируют in vivo и
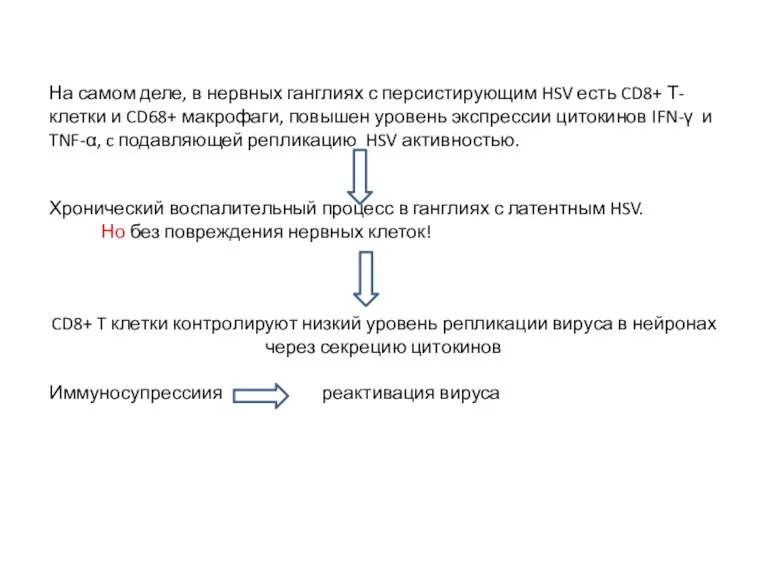
- 11. На самом деле, в нервных ганглиях с персистирующим HSV есть CD8+ Т-клетки и CD68+ макрофаги, повышен
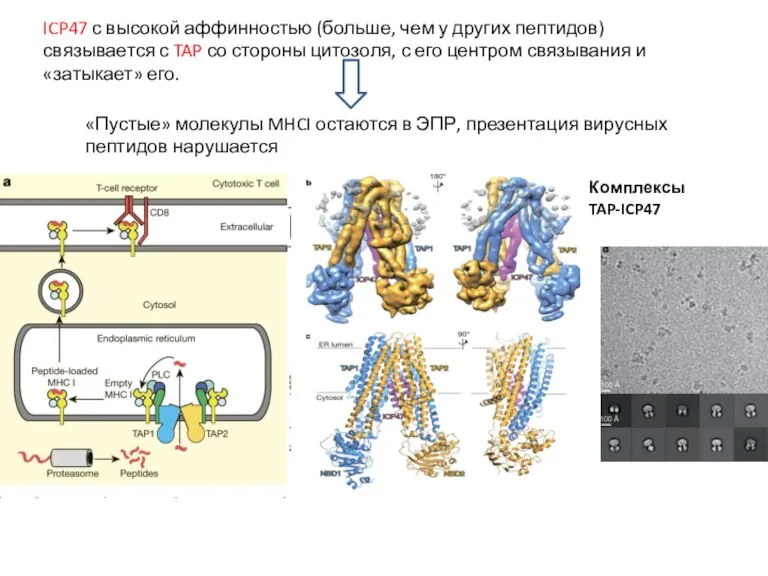
- 12. Каковы молекулярные механизмы избегания вирусом HSV нормального иммунного ответа в клетках эпителия? M. L. Oldham, R.K.
- 13. ICP47 с высокой аффинностью (больше, чем у других пептидов) связывается с TAP со стороны цитозоля, с
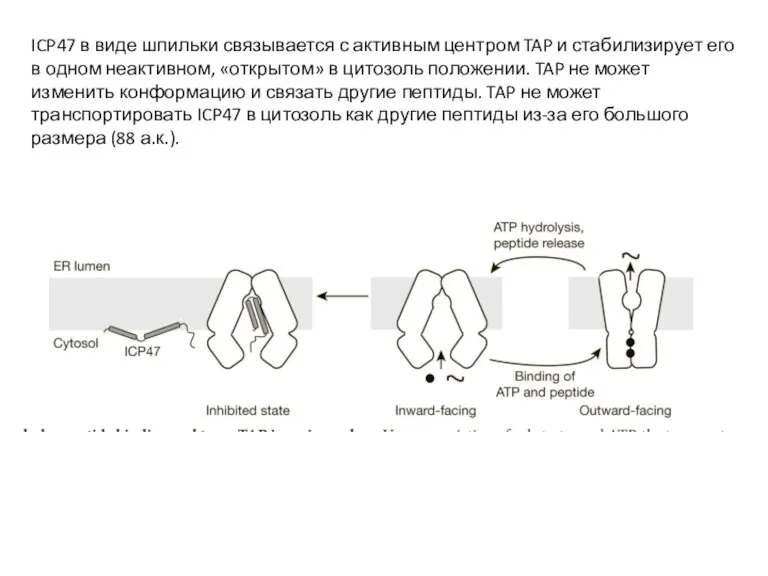
- 14. ICP47 в виде шпильки связывается с активным центром TAP и стабилизирует его в одном неактивном, «открытом»
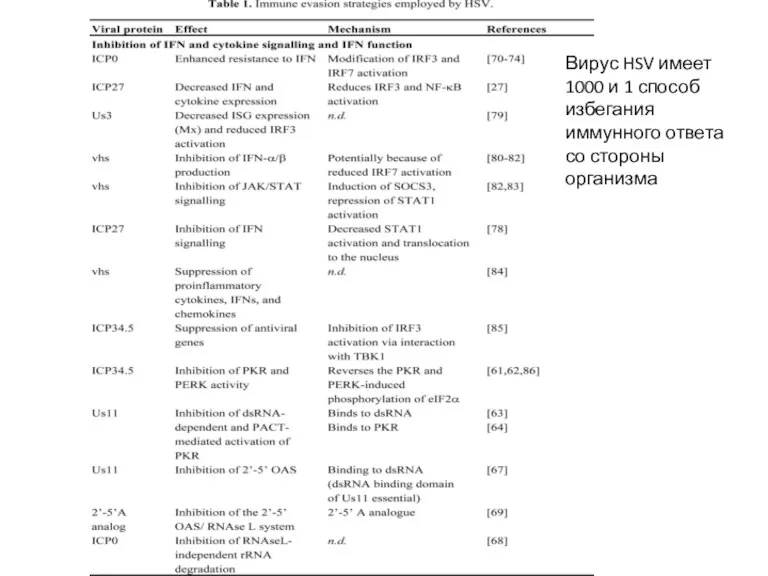
- 15. Вирус HSV имеет 1000 и 1 способ избегания иммунного ответа со стороны организма
- 18. Epstein-Barr virus EBV Инфицирует В-клетки через СD21 и MHCII. Первичная инфекция – или бессимптомно или инфекционный
- 19. Некоторые патогены устойчивы к защитным механизмам хозяина или используют его в своих собственных целях Micobacterium tuberculosis
- 20. Врожденные иммунодефициты (более 100) –дефекты в генах, участвующих в контроле иммунного ответа Общая особенность - в
- 21. Дефекты в развитии Т- и В-клеток, вызывающие иммунодефициты. Дефекты Т-клеток – самые тяжелые SCID – severe
- 22. Синдром приобретенного иммунодефицита СПИД/AIDS подавление иммунного ответа, вызванное инфекцией HIV (human immunodeficiency virus) 1981 – первое
- 23. Пути попадания HIV в организм человека: Через слизистую половых путей – основной путь С кровью (через
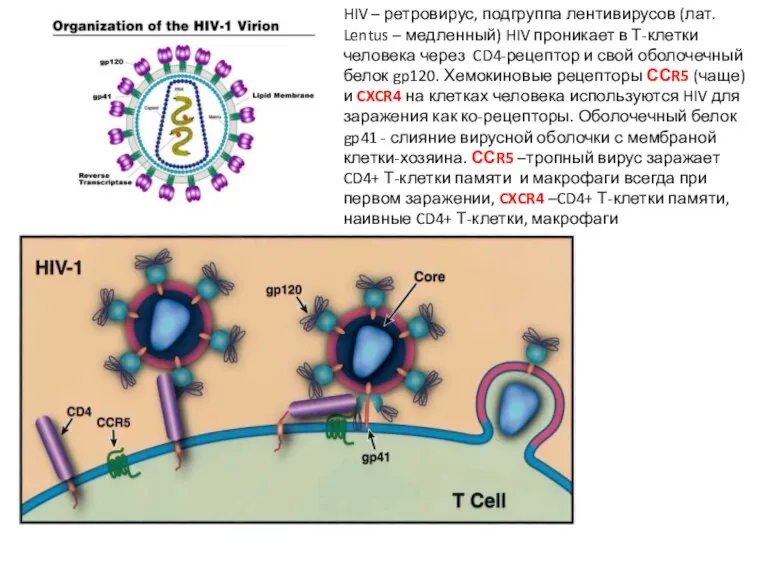
- 24. HIV – ретровирус, подгруппа лентивирусов (лат. Lentus – медленный) HIV проникает в Т-клетки человека через CD4-рецептор
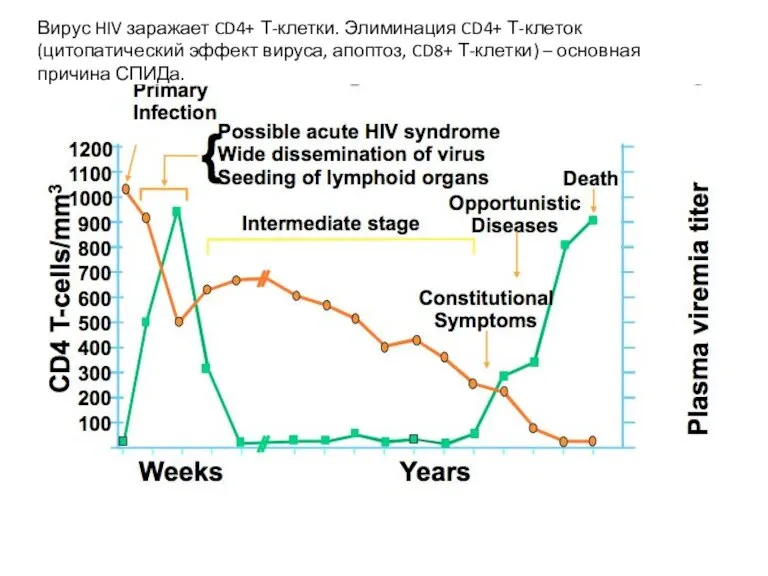
- 25. Вирус HIV заражает CD4+ Т-клетки. Элиминация CD4+ Т-клеток (цитопатический эффект вируса, апоптоз, CD8+ Т-клетки) – основная
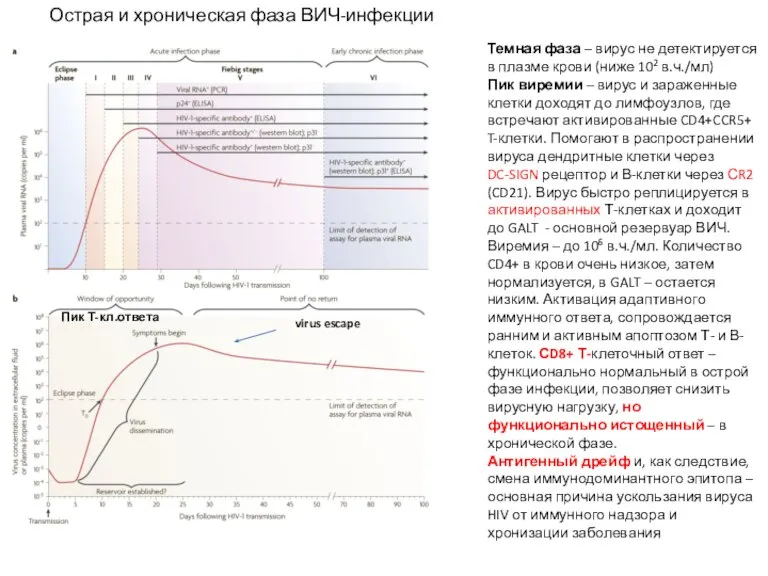
- 26. Темная фаза – вирус не детектируется в плазме крови (ниже 102 в.ч./мл) Пик виремии – вирус
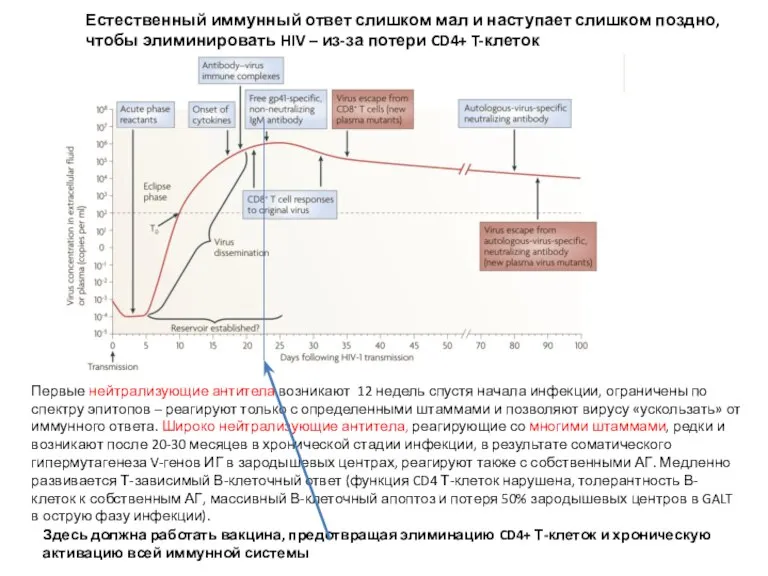
- 27. Первые нейтрализующие антитела возникают 12 недель спустя начала инфекции, ограничены по спектру эпитопов – реагируют только
- 28. Вакцина против ВИЧ должна обеспечивать присутствие широко нейтрализующих антител в плазме и слизистых
- 29. Антигенный дрейф и функциональное истощение CTLs – основные причины неспособности иммунной системы человека элиминировать ВИЧ
- 30. ~ через 30 дней вирус-специфические ЦТЛ Env and Nef, whereas Gag-specific and Pol-specific responses tend to
- 31. Хроническая фаза ВИЧ-инфекции Результат хронической антигенной стимуляции CD8+ T-клеток и, как следствие, повышенной экспрессии ингибирующих рецепторов
- 32. PD1 - №1 в функциональном истощении Т-клеток, основной регулятор. ITIM-содержащий рецептор, экспрессия всегда повышается при активации
- 33. Около 1% ВИЧ-зараженных – «элит-контроллеры» или «элит-нон-прогрессоры» или «элит-супрессоры». Без ретровирусной терапии – вирусная нагрузка не
- 36. ССR5del Это теперь не зазорно! 2008. Timothy Brown, also known as the «Berlin patient» 1995 –
- 38. Скачать презентацию
Слайд 2Иммунодефициты
Первичные Вторичные
(врожденные) (приобретенные)
Неспособность высшего организма противостоять инфекции
Избегание патогеном действия иммунной системы
Иммунодефициты
Первичные Вторичные
(врожденные) (приобретенные)
Неспособность высшего организма противостоять инфекции
Избегание патогеном действия иммунной системы
Слайд 3Многие инфекционные агенты существуют в нескольких антигенных вариантах.
Streptococcus pneumoniae – 84
Многие инфекционные агенты существуют в нескольких антигенных вариантах.
Streptococcus pneumoniae – 84

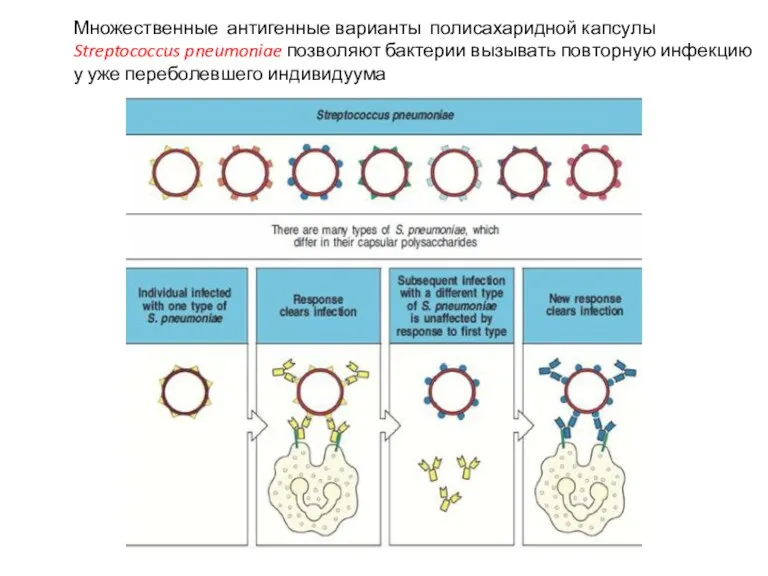
Слайд 4Множественные антигенные варианты полисахаридной капсулы Streptococcus pneumoniae позволяют бактерии вызывать повторную инфекцию
Множественные антигенные варианты полисахаридной капсулы Streptococcus pneumoniae позволяют бактерии вызывать повторную инфекцию
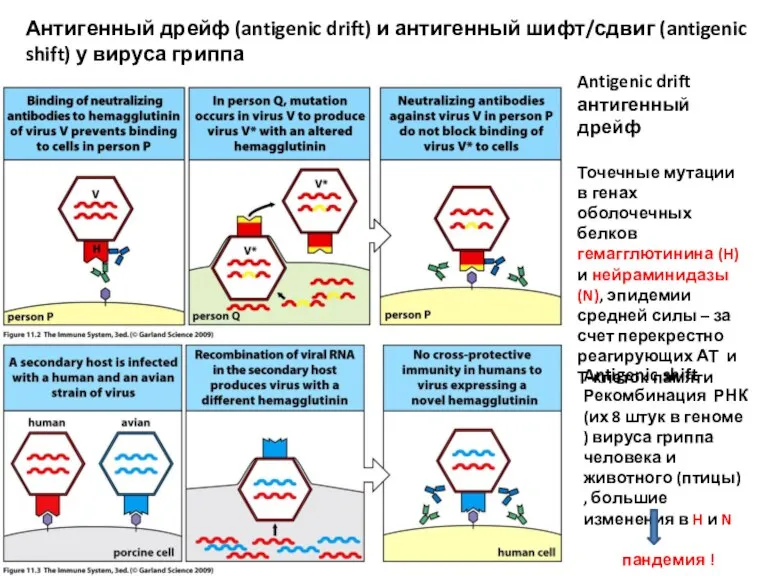
Слайд 5Антигенный дрейф (antigenic drift) и антигенный шифт/сдвиг (antigenic shift) у вируса гриппа
Антигенный дрейф (antigenic drift) и антигенный шифт/сдвиг (antigenic shift) у вируса гриппа
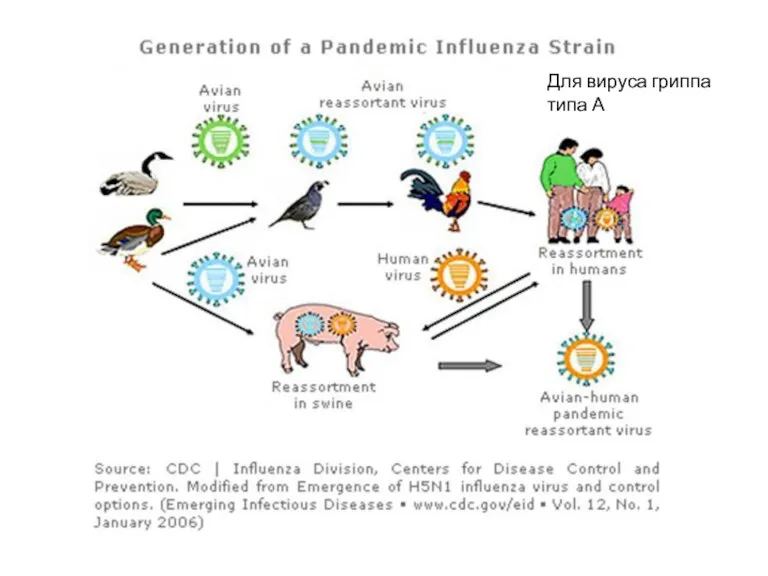
Слайд 6Для вируса гриппа типа А
Для вируса гриппа типа А
Слайд 7Смена подтипов вируса гриппа А и связанные с ней пандемии и эпидемии
Смена подтипов вируса гриппа А и связанные с ней пандемии и эпидемии
Слайд 8Запрограммированные перестройки в ДНК патогена
Trypanosoma brucei , простейшее, переносчик – муха цеце, у
Запрограммированные перестройки в ДНК патогена
Trypanosoma brucei , простейшее, переносчик – муха цеце, у
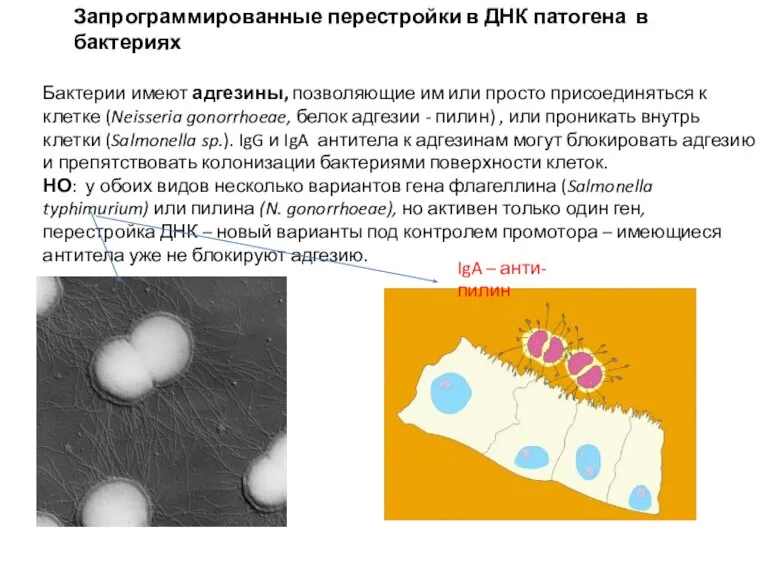
Слайд 9Бактерии имеют адгезины, позволяющие им или просто присоединяться к клетке (Neisseria gonorrhoeae,
Бактерии имеют адгезины, позволяющие им или просто присоединяться к клетке (Neisseria gonorrhoeae,
Слайд 10Некоторые вирусы могут «исчезать» из видимости иммунной системы в латентный период -
Некоторые вирусы могут «исчезать» из видимости иммунной системы в латентный период -
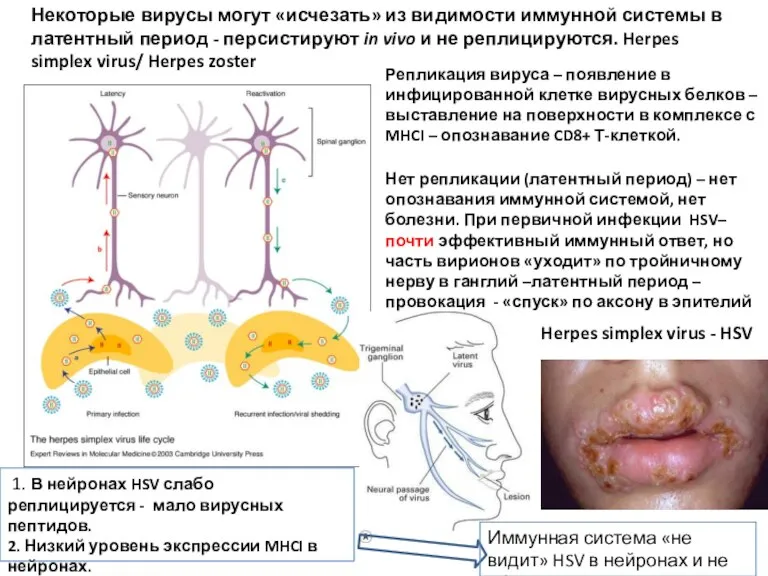
Слайд 11На самом деле, в нервных ганглиях с персистирующим HSV есть CD8+ Т-клетки
На самом деле, в нервных ганглиях с персистирующим HSV есть CD8+ Т-клетки
Слайд 12Каковы молекулярные механизмы избегания вирусом HSV нормального иммунного
ответа в клетках эпителия?
Каковы молекулярные механизмы избегания вирусом HSV нормального иммунного
ответа в клетках эпителия?

Слайд 13ICP47 с высокой аффинностью (больше, чем у других пептидов) связывается с TAP
ICP47 с высокой аффинностью (больше, чем у других пептидов) связывается с TAP
Слайд 14ICP47 в виде шпильки связывается с активным центром TAP и стабилизирует его
ICP47 в виде шпильки связывается с активным центром TAP и стабилизирует его
Слайд 15Вирус HSV имеет 1000 и 1 способ избегания иммунного ответа со стороны
Вирус HSV имеет 1000 и 1 способ избегания иммунного ответа со стороны
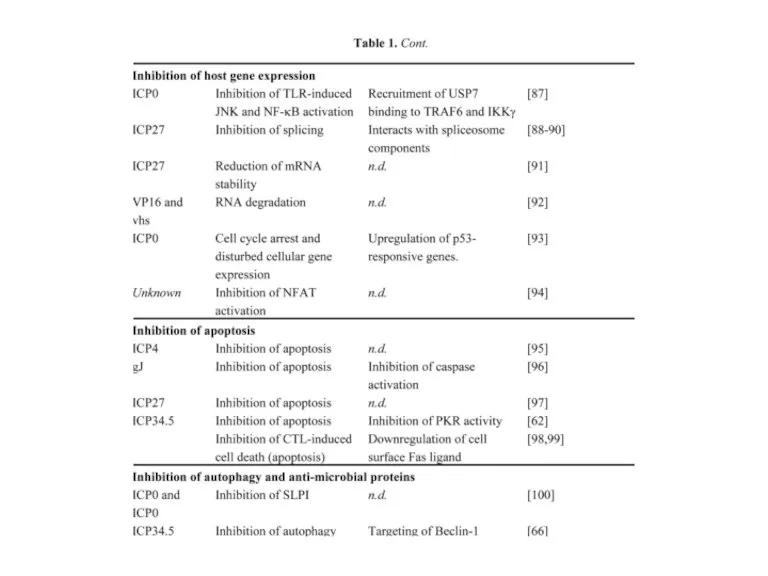
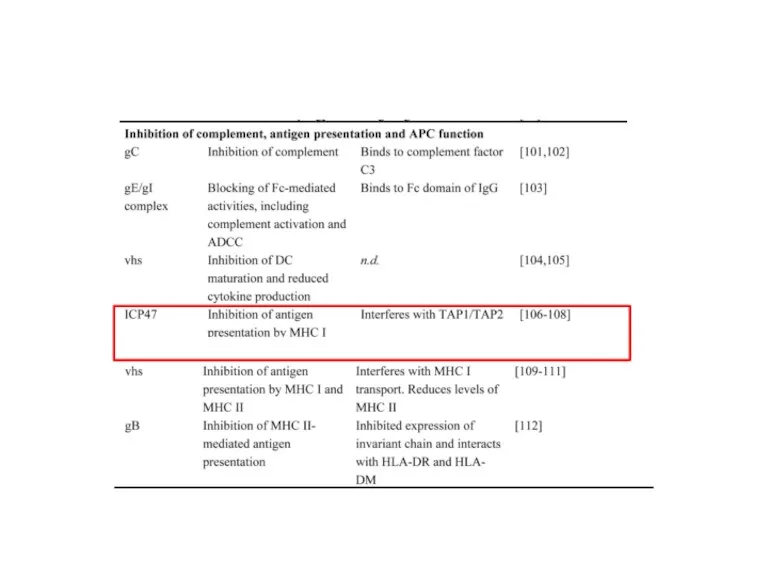
Слайд 18 Epstein-Barr virus EBV
Инфицирует В-клетки через СD21 и MHCII.
Первичная инфекция – или
Epstein-Barr virus EBV
Инфицирует В-клетки через СD21 и MHCII.
Первичная инфекция – или

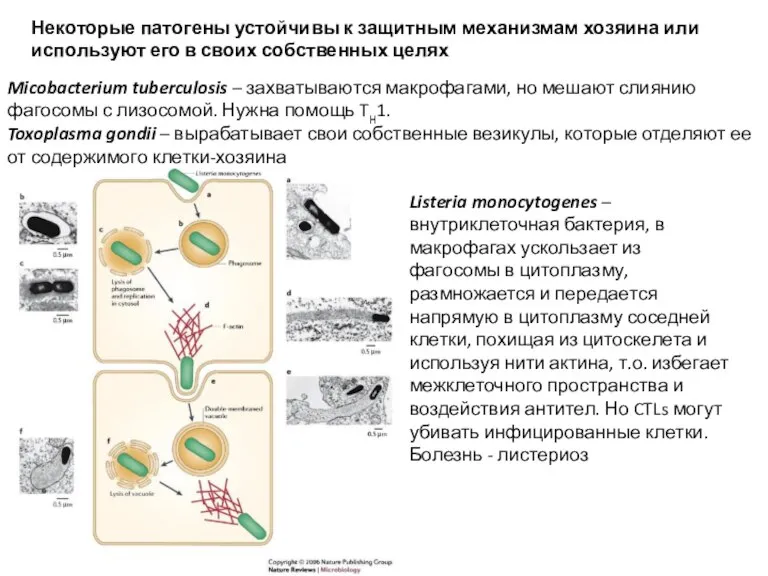
Слайд 19Некоторые патогены устойчивы к защитным механизмам хозяина или используют его в своих
Некоторые патогены устойчивы к защитным механизмам хозяина или используют его в своих
Слайд 20Врожденные иммунодефициты (более 100) –дефекты в генах, участвующих в контроле иммунного ответа
Общая
Врожденные иммунодефициты (более 100) –дефекты в генах, участвующих в контроле иммунного ответа
Общая

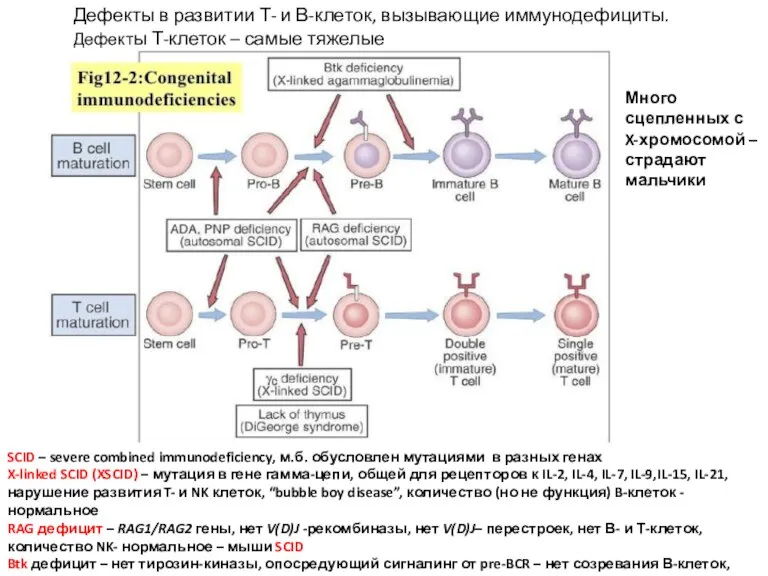
Слайд 21Дефекты в развитии Т- и В-клеток, вызывающие иммунодефициты. Дефекты Т-клеток – самые
Дефекты в развитии Т- и В-клеток, вызывающие иммунодефициты. Дефекты Т-клеток – самые
Слайд 22 Синдром приобретенного иммунодефицита СПИД/AIDS
подавление иммунного ответа, вызванное инфекцией HIV (human immunodeficiency
Синдром приобретенного иммунодефицита СПИД/AIDS
подавление иммунного ответа, вызванное инфекцией HIV (human immunodeficiency

Слайд 23 Пути попадания HIV в организм человека:
Через слизистую половых путей – основной
Пути попадания HIV в организм человека:
Через слизистую половых путей – основной

Слайд 24HIV – ретровирус, подгруппа лентивирусов (лат. Lentus – медленный) HIV проникает в
HIV – ретровирус, подгруппа лентивирусов (лат. Lentus – медленный) HIV проникает в
Слайд 25Вирус HIV заражает CD4+ Т-клетки. Элиминация CD4+ Т-клеток (цитопатический эффект вируса, апоптоз,
Вирус HIV заражает CD4+ Т-клетки. Элиминация CD4+ Т-клеток (цитопатический эффект вируса, апоптоз,
Слайд 26Темная фаза – вирус не детектируется в плазме крови (ниже 102 в.ч./мл)
Пик
Темная фаза – вирус не детектируется в плазме крови (ниже 102 в.ч./мл)
Пик
Слайд 27Первые нейтрализующие антитела возникают 12 недель спустя начала инфекции, ограничены по спектру
Первые нейтрализующие антитела возникают 12 недель спустя начала инфекции, ограничены по спектру
Слайд 28Вакцина против ВИЧ должна обеспечивать присутствие широко нейтрализующих антител в плазме и
Вакцина против ВИЧ должна обеспечивать присутствие широко нейтрализующих антител в плазме и

Слайд 29Антигенный дрейф и функциональное истощение CTLs – основные причины неспособности иммунной системы
Антигенный дрейф и функциональное истощение CTLs – основные причины неспособности иммунной системы

Слайд 30~ через 30 дней вирус-специфические ЦТЛ
Env and Nef, whereas Gag-specific and
~ через 30 дней вирус-специфические ЦТЛ
Env and Nef, whereas Gag-specific and
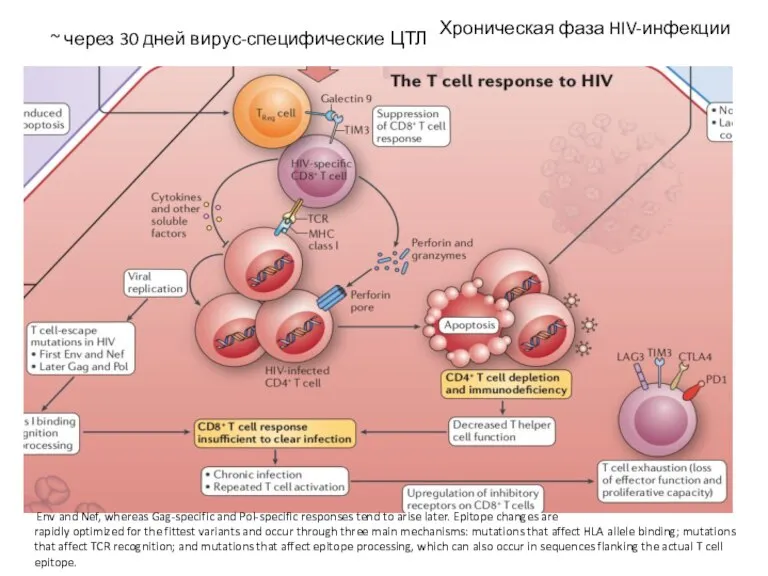
Слайд 31 Хроническая фаза ВИЧ-инфекции
Результат хронической антигенной стимуляции CD8+ T-клеток и, как следствие,
Хроническая фаза ВИЧ-инфекции
Результат хронической антигенной стимуляции CD8+ T-клеток и, как следствие,

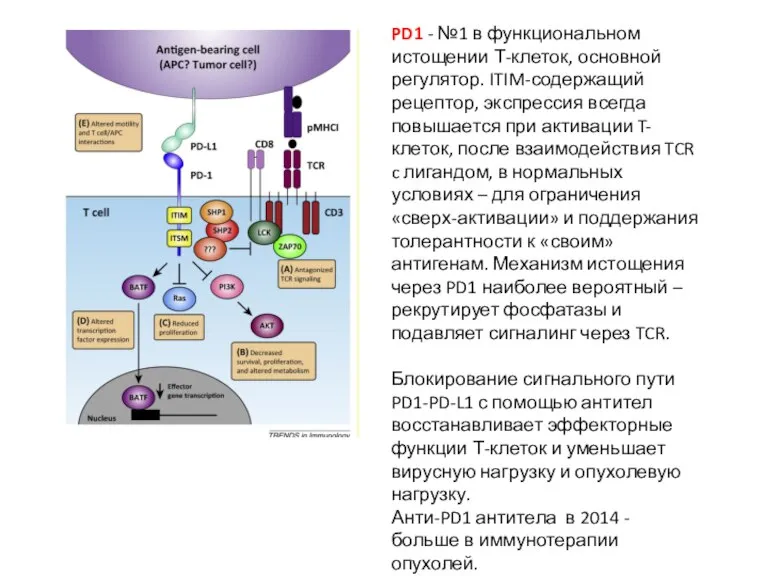
Слайд 32PD1 - №1 в функциональном истощении Т-клеток, основной регулятор. ITIM-содержащий рецептор, экспрессия
PD1 - №1 в функциональном истощении Т-клеток, основной регулятор. ITIM-содержащий рецептор, экспрессия
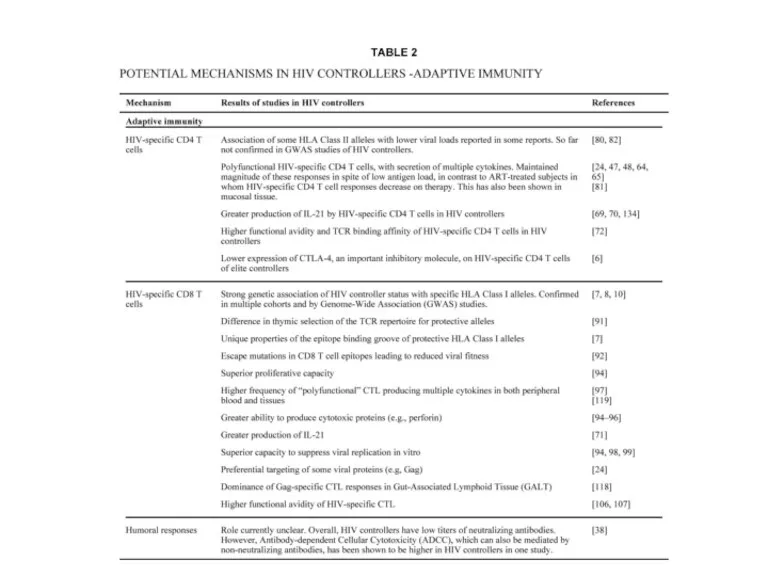
Слайд 33Около 1% ВИЧ-зараженных – «элит-контроллеры» или «элит-нон-прогрессоры» или «элит-супрессоры».
Без ретровирусной терапии –
Около 1% ВИЧ-зараженных – «элит-контроллеры» или «элит-нон-прогрессоры» или «элит-супрессоры».
Без ретровирусной терапии –


Слайд 36ССR5del
Это теперь не зазорно!
2008. Timothy Brown, also known as the «Berlin patient»
1995
ССR5del
Это теперь не зазорно!
2008. Timothy Brown, also known as the «Berlin patient»
1995

 Су алмасуы патофизиологиясы
Су алмасуы патофизиологиясы Алгоритм, диагностика и оказание неотложной помощи при синдроме длительного сдавления
Алгоритм, диагностика и оказание неотложной помощи при синдроме длительного сдавления Профилактика гриппа
Профилактика гриппа Что такое ферменты?
Что такое ферменты? Травма живота. Лекция
Травма живота. Лекция Хирургическое лечение глубоких ожогов у детей
Хирургическое лечение глубоких ожогов у детей Анатомо-физиологические особенности детей дошкольного возраста
Анатомо-физиологические особенности детей дошкольного возраста Плоскостопия можно избежать
Плоскостопия можно избежать Практикум по ЭКГ
Практикум по ЭКГ Психопатологическая семиотика и синдромология. Первая помощь при эпилепсии
Психопатологическая семиотика и синдромология. Первая помощь при эпилепсии Понятие, признаки и виды вспомогательных репродуктивных технологий. Нормативно-правовое регулирование
Понятие, признаки и виды вспомогательных репродуктивных технологий. Нормативно-правовое регулирование Псориатическая артропатия
Псориатическая артропатия Коронавирус
Коронавирус Заболевания ОДА. Остеохондроз позвоночного столба
Заболевания ОДА. Остеохондроз позвоночного столба Силикоз
Силикоз Репродуктивное здоровье студентов
Репродуктивное здоровье студентов Стратификация (градация) стадий ХБП по уровням СКФ
Стратификация (градация) стадий ХБП по уровням СКФ Heart failure slideset
Heart failure slideset Разрыв мениска
Разрыв мениска Сосудистые заболевания головного мозга. Лекция 3. 1 часть
Сосудистые заболевания головного мозга. Лекция 3. 1 часть Геронтология
Геронтология От чего мы иногда болеем
От чего мы иногда болеем Курация больных ГЭРБ (гастроэзофагеальной рефлюксной болезни)
Курация больных ГЭРБ (гастроэзофагеальной рефлюксной болезни) Роль наддесневых зубных камней в развитии пародонтоза
Роль наддесневых зубных камней в развитии пародонтоза Гнойничковые заболевания кожи
Гнойничковые заболевания кожи Основы патологии
Основы патологии Первая медицинская помощь при острой сердечной недостаточности и инсульте
Первая медицинская помощь при острой сердечной недостаточности и инсульте Грибковые заболевания кожи и волос
Грибковые заболевания кожи и волос