- Разнообразие иммунных реакций беспозвоночных

Содержание
- 2. Иммуноциты беспозвоночных
- 4. Строение гидры
- 5. Эпителиальные клетки гидроидных способны к фагоцитозу
- 6. У книдарий есть собственные TLR- и NLR - рецепторы
- 7. Антимикробные пептиды книдарий
- 8. Странный белок дождевого червя CCF Белок жидкости целомической полости массой 42 кДа, распознаёт (?) О-антиген ЛПС,
- 9. Хлорагогенные клетки, малые целомоциты и амёбоциты кольчатых червей
- 10. Антимикробные пептиды пиявки
- 11. Строение FREP белков улитки Biomphalaria glabrata
- 12. Улитки против трематод
- 13. Различия между чувствительными и устойчивыми улитками
- 14. Гипотеза черной королевы
- 15. Половое размножение помогает устойчивости к паразитам на популяционном уровне
- 16. Euprymna scolopes
- 17. Строение светового органа эупримны
- 18. Vibrio fischeri прикрепляются к ресничному эпителию светового органа каракатицы Интенсивность свечения зависит от чувства кворума
- 19. Каракатица регулирует количество симбионтов в световом органе
- 20. Заселение светового органа каракатицы после рождения
- 21. Модель колонизации
- 22. Деградация ресничного эпителия светового органа идет в ответ на MAMP Vibrio fischeri
- 23. Общее строении нематоды 1031 клетка у самца, 959 клеток у гермафродита и ни одного иммуноцита!
- 24. Патогены нематод и пути их проникновения
- 25. Грибковые инфекции нематод
- 26. МАР-киназный каскад как сигнал о повреждении покровов Гидроксифенилакриловая кислота
- 27. Иммунная система нематод
- 28. Сигнальные пути, вовлеченные в иммунный ответ нематоды, еще не известны полностью
- 29. Различие сигнальных путей
- 30. а
- 31. Микробиота нематоды и старение
- 32. Общая схема строения морского ежа
- 33. Характеристика генов иммунной системы иглокожих и хордовых
- 34. Аксиальный орган иглокожих
- 35. «Формула» целомоцитов морского ежа
- 36. Целомоциты морского ежа:
- 37. а
- 38. Полигональные и дисковидные фагоциты отличаются организацией цитоскелета
- 39. Рецепторные белки иглокожих
- 40. Рецепторы целомоцита асцидии
- 41. Гены VCBP у ланцетника
- 42. «Тренированный» врождённый иммунитет – аналог адаптивного?
- 44. Граница между врожденным и адаптивным иммунитетом
- 45. Направления эволюции иммунных рецепторов
- 47. Скачать презентацию

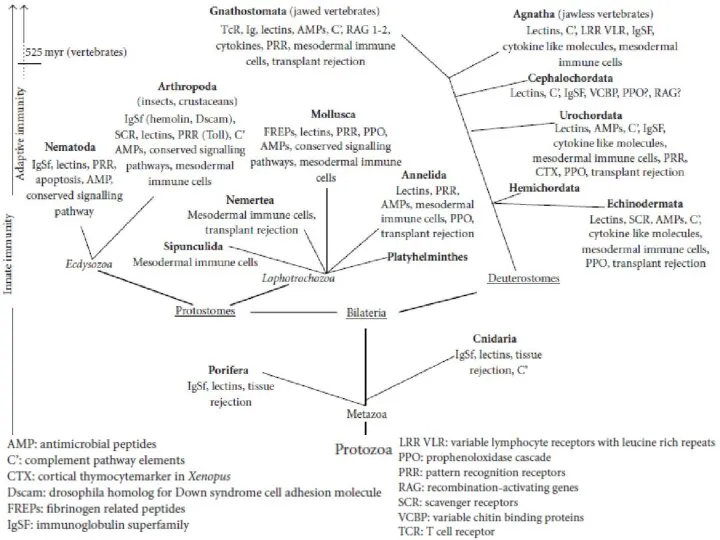
Слайд 4Строение гидры
Строение гидры
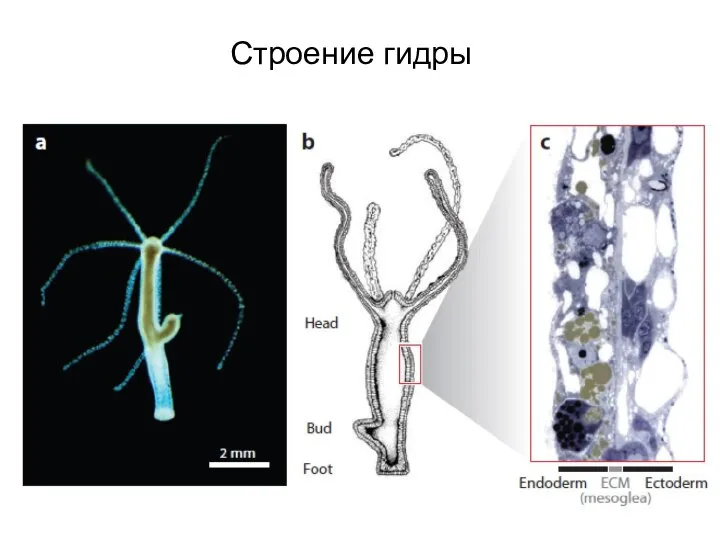
Слайд 5Эпителиальные клетки гидроидных способны к фагоцитозу
Эпителиальные клетки гидроидных способны к фагоцитозу
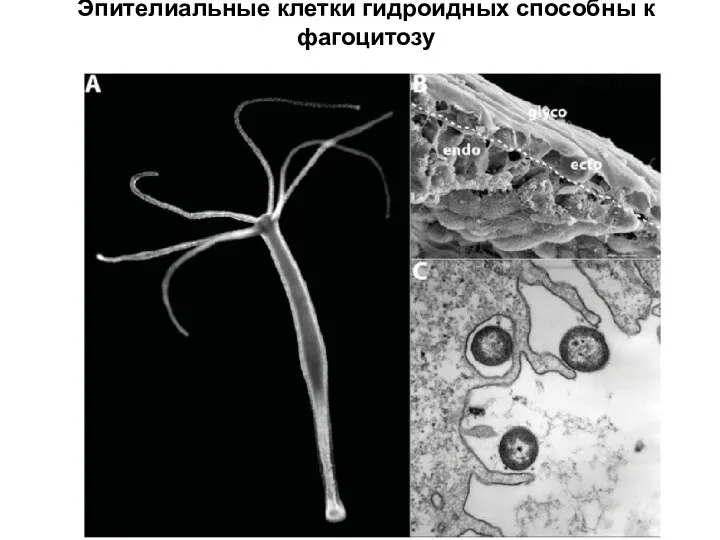
Слайд 6У книдарий есть собственные TLR- и NLR - рецепторы
У книдарий есть собственные TLR- и NLR - рецепторы
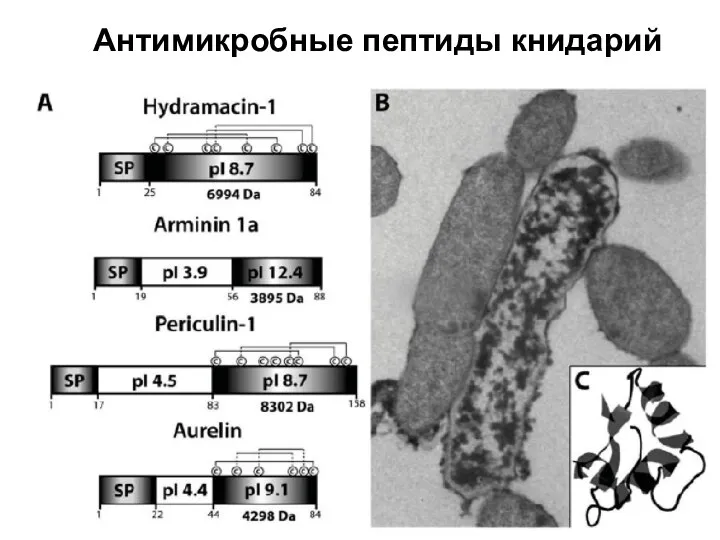
Слайд 7Антимикробные пептиды книдарий
Антимикробные пептиды книдарий
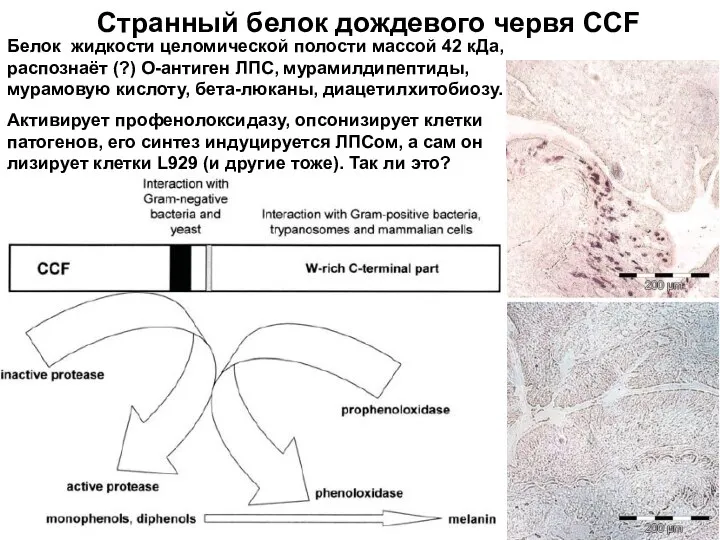
Слайд 8Странный белок дождевого червя CCF
Белок жидкости целомической полости массой 42 кДа, распознаёт
Странный белок дождевого червя CCF
Белок жидкости целомической полости массой 42 кДа, распознаёт
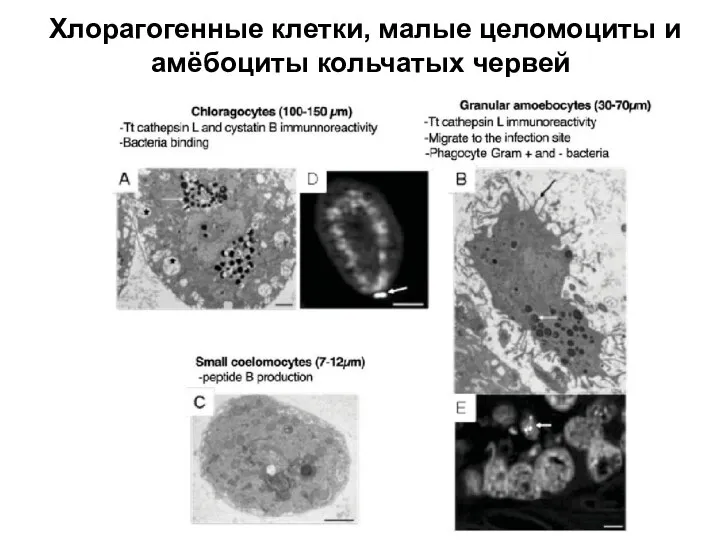
Слайд 9Хлорагогенные клетки, малые целомоциты и амёбоциты кольчатых червей
Хлорагогенные клетки, малые целомоциты и амёбоциты кольчатых червей
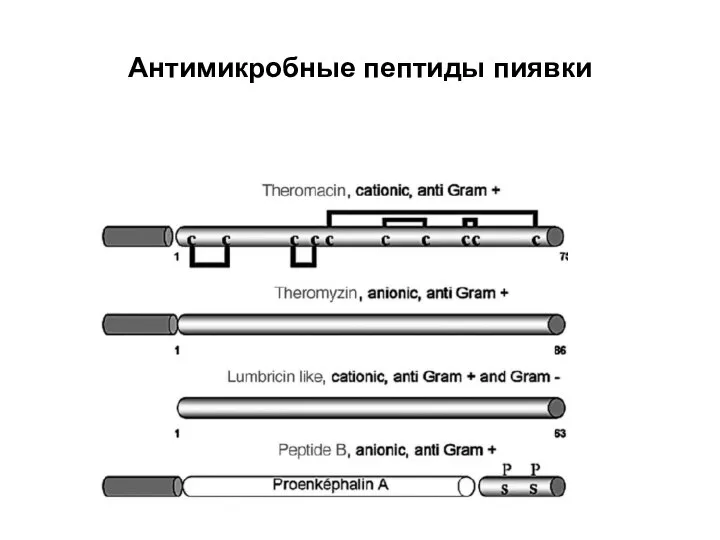
Слайд 10Антимикробные пептиды пиявки
Антимикробные пептиды пиявки
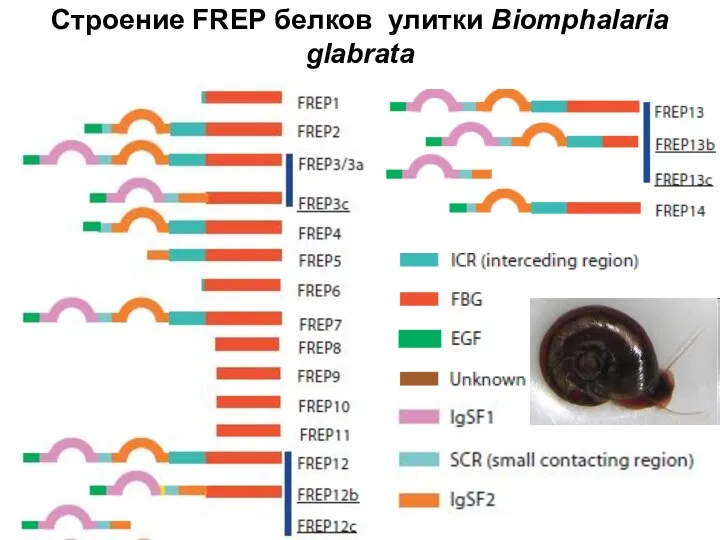
Слайд 11Строение FREP белков улитки Biomphalaria
glabrata
Строение FREP белков улитки Biomphalaria
glabrata
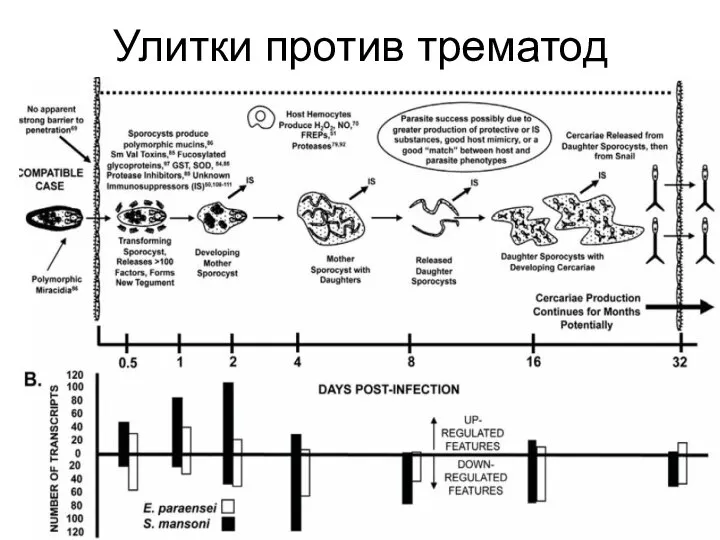
Слайд 12Улитки против трематод
Улитки против трематод
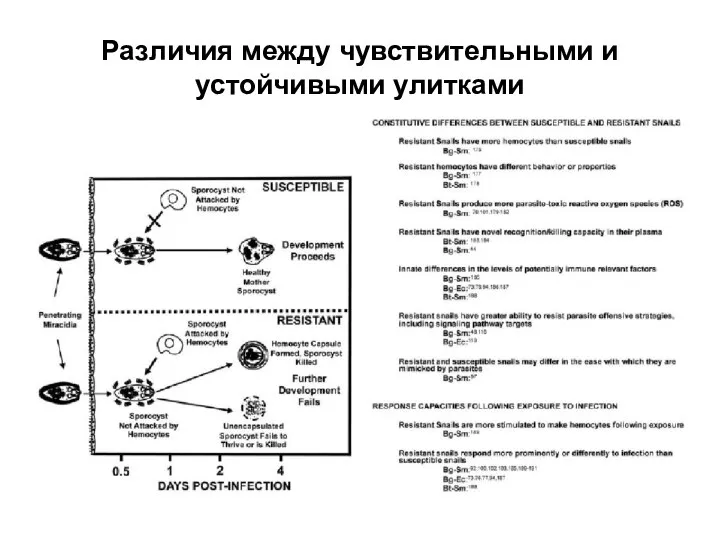
Слайд 13Различия между чувствительными и устойчивыми улитками
Различия между чувствительными и устойчивыми улитками
Слайд 14Гипотеза черной королевы
Гипотеза черной королевы

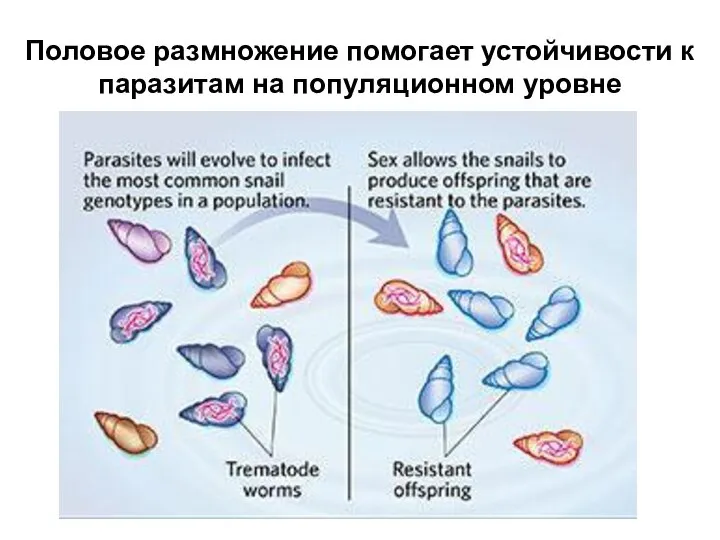
Слайд 15Половое размножение помогает устойчивости к паразитам на популяционном уровне
Половое размножение помогает устойчивости к паразитам на популяционном уровне
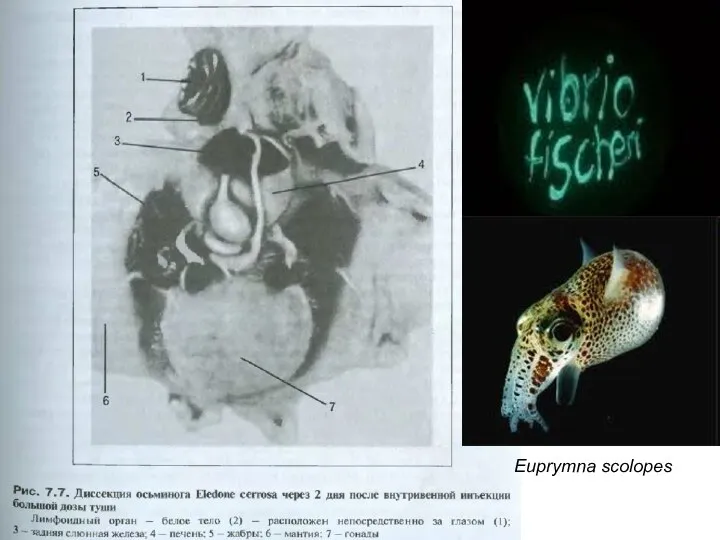
Слайд 16Euprymna scolopes
Euprymna scolopes
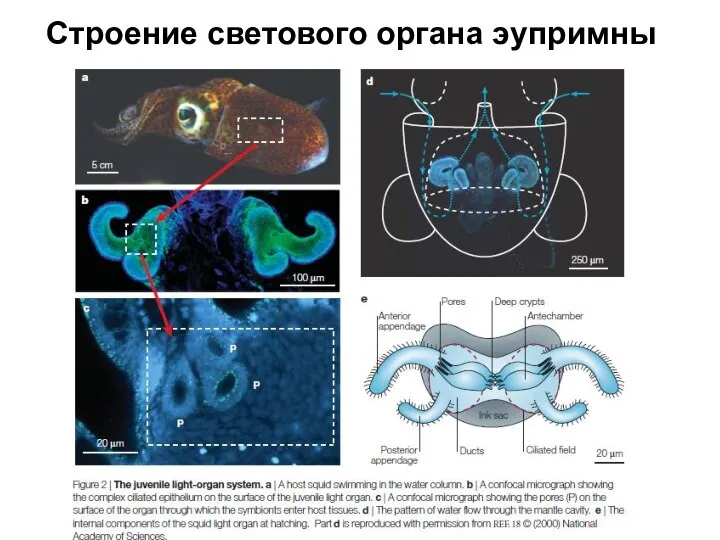
Слайд 17Строение светового органа эупримны
Строение светового органа эупримны
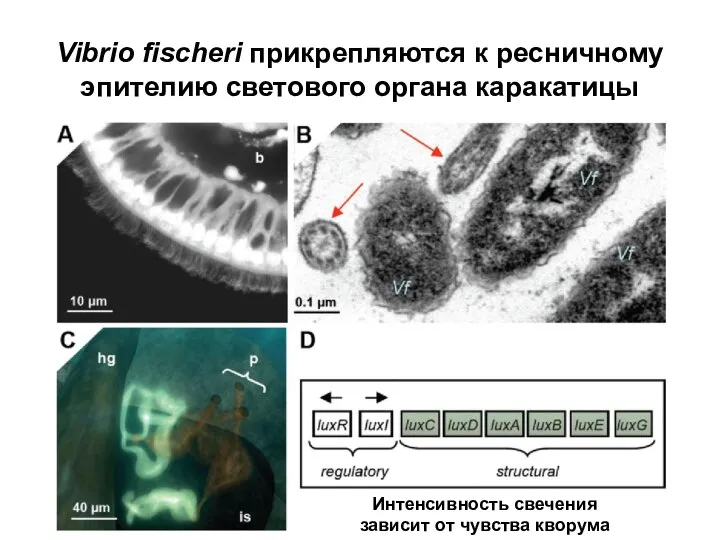
Слайд 18Vibrio fischeri прикрепляются к ресничному эпителию светового органа каракатицы
Интенсивность свечения зависит от
Vibrio fischeri прикрепляются к ресничному эпителию светового органа каракатицы
Интенсивность свечения зависит от
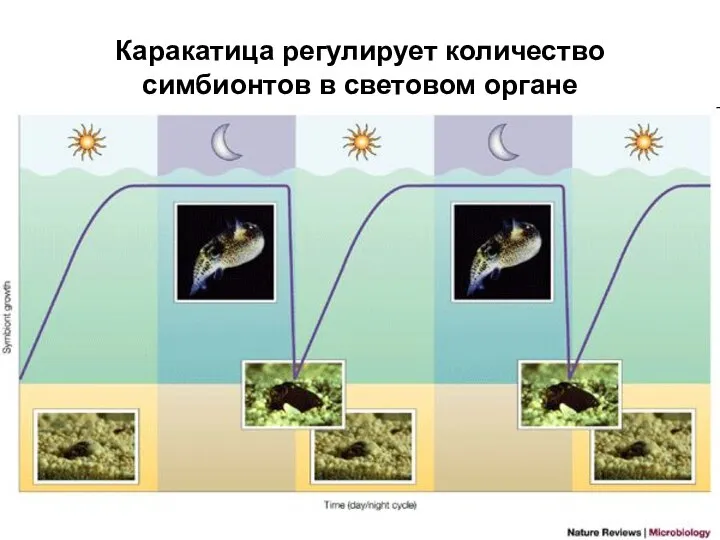
Слайд 19Каракатица регулирует количество симбионтов в световом органе
Каракатица регулирует количество симбионтов в световом органе
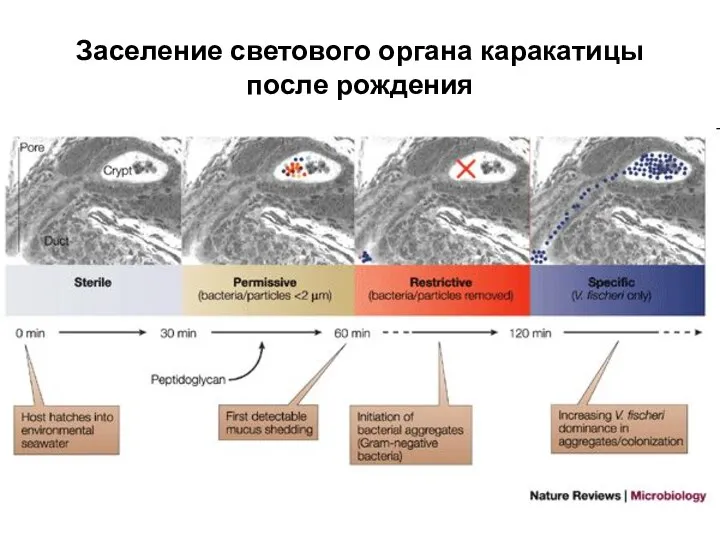
Слайд 20Заселение светового органа каракатицы после рождения
Заселение светового органа каракатицы после рождения
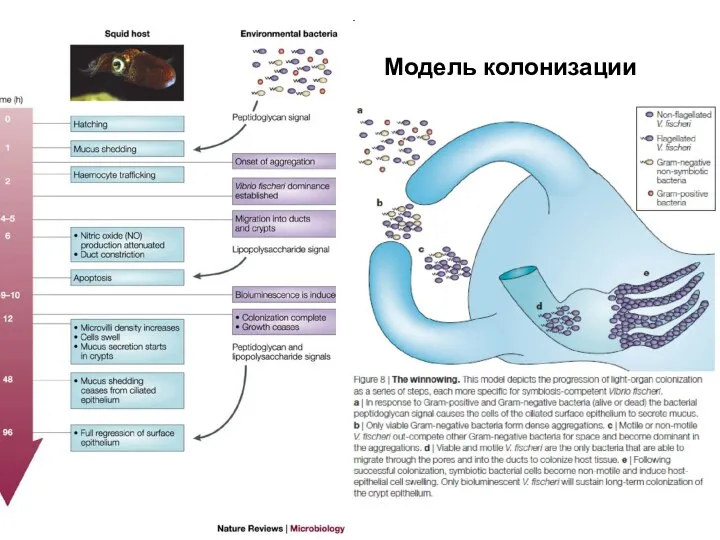
Слайд 21Модель колонизации
Модель колонизации
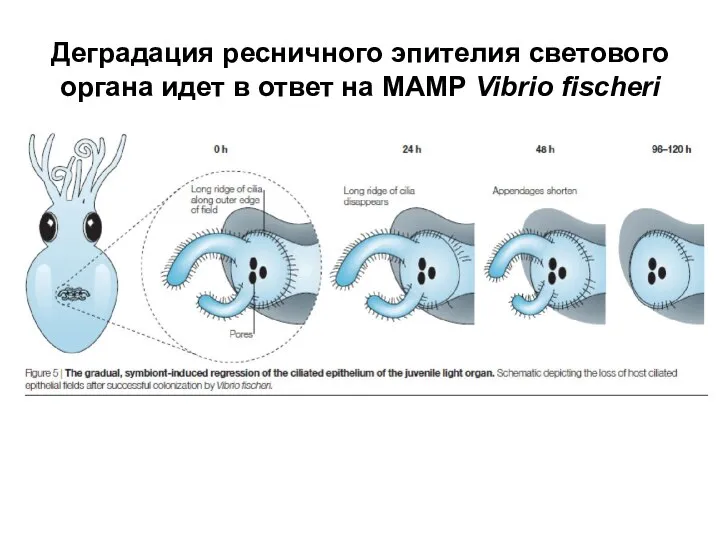
Слайд 22Деградация ресничного эпителия светового органа идет в ответ на MAMP Vibrio fischeri
Деградация ресничного эпителия светового органа идет в ответ на MAMP Vibrio fischeri
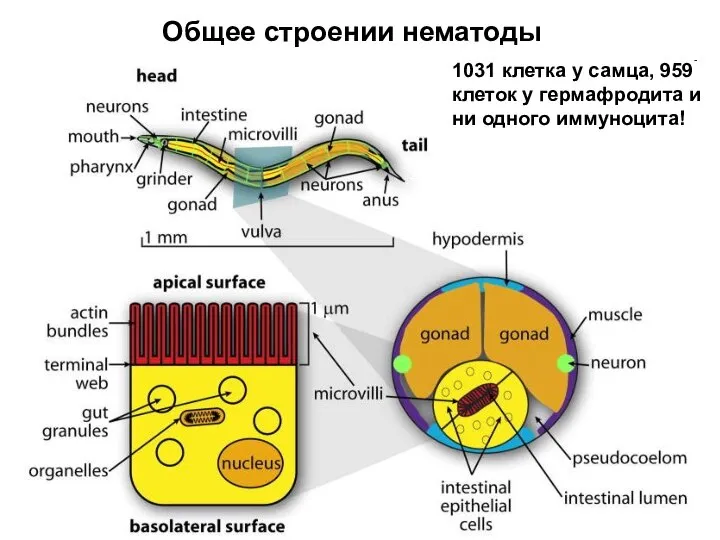
Слайд 23Общее строении нематоды
1031 клетка у самца, 959 клеток у гермафродита и ни
Общее строении нематоды
1031 клетка у самца, 959 клеток у гермафродита и ни
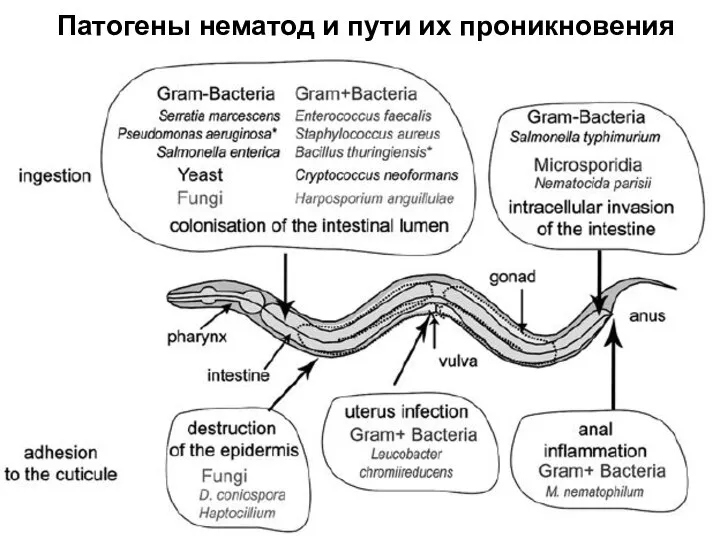
Слайд 24Патогены нематод и пути их проникновения
Патогены нематод и пути их проникновения
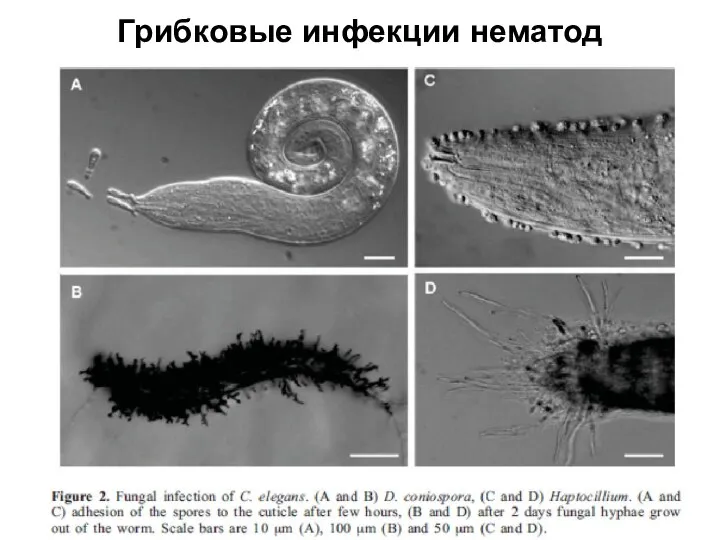
Слайд 25Грибковые инфекции нематод
Грибковые инфекции нематод
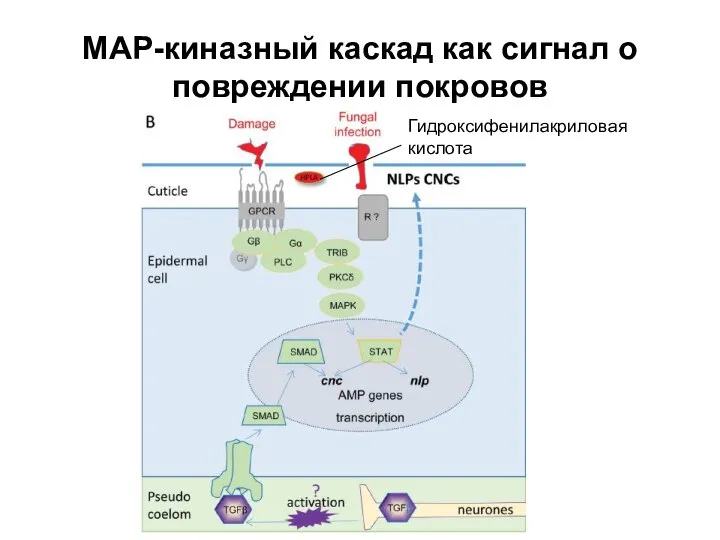
Слайд 26МАР-киназный каскад как сигнал о повреждении покровов
Гидроксифенилакриловая кислота
МАР-киназный каскад как сигнал о повреждении покровов
Гидроксифенилакриловая кислота
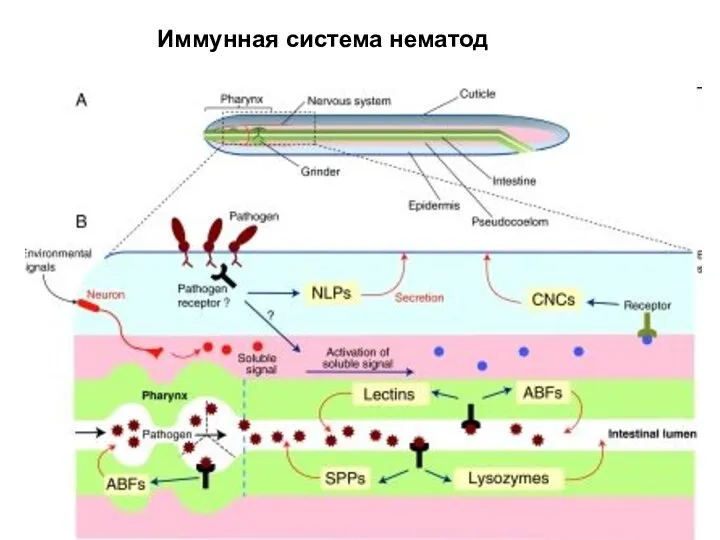
Слайд 27Иммунная система нематод
Иммунная система нематод
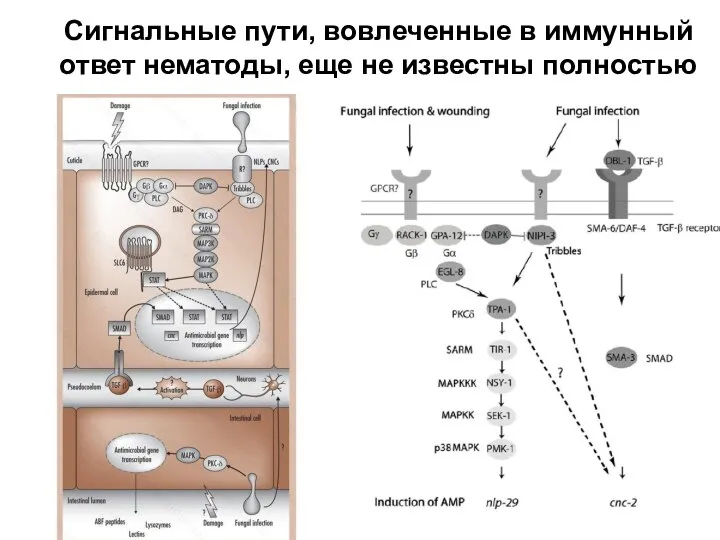
Слайд 28Сигнальные пути, вовлеченные в иммунный ответ нематоды, еще не известны полностью
Сигнальные пути, вовлеченные в иммунный ответ нематоды, еще не известны полностью
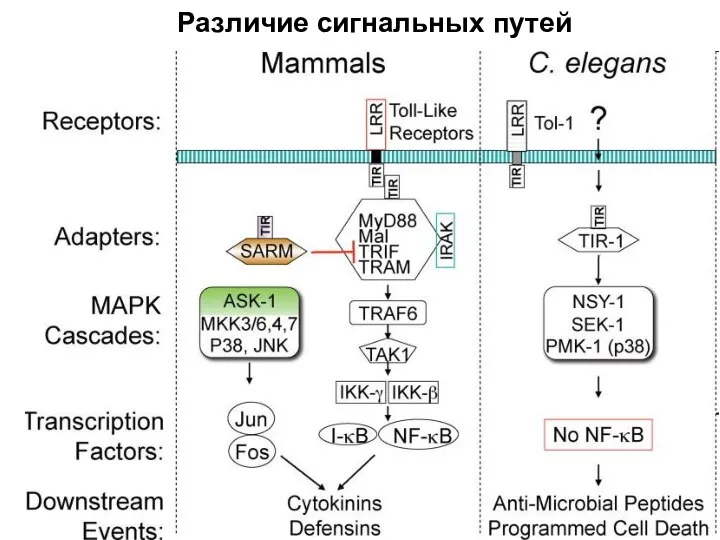
Слайд 29Различие сигнальных путей
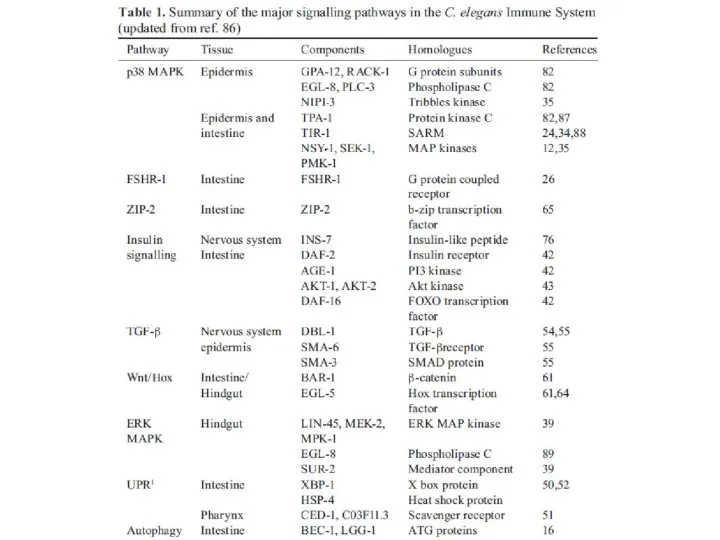
Различие сигнальных путей
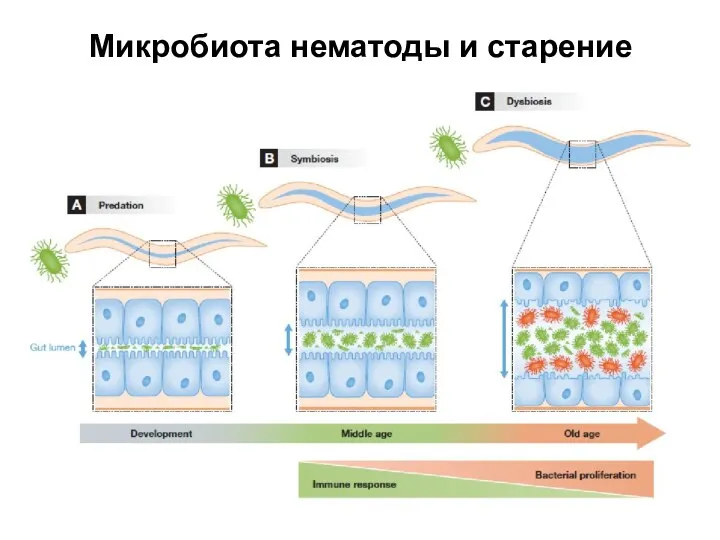
Слайд 31Микробиота нематоды и старение
Микробиота нематоды и старение
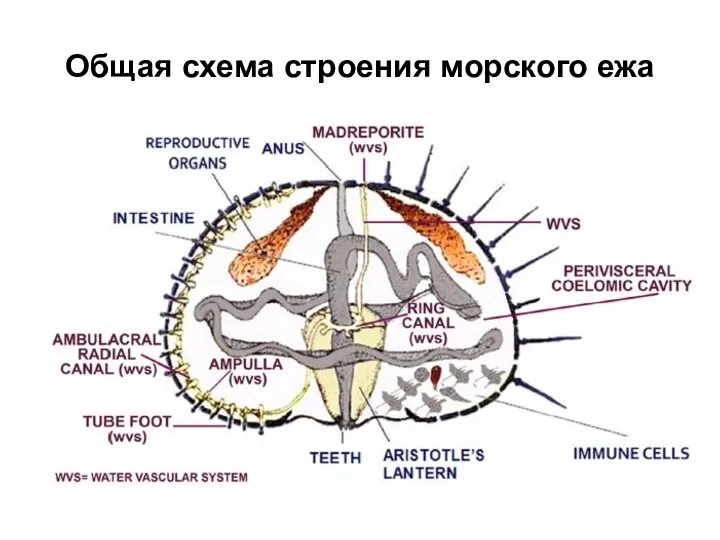
Слайд 32Общая схема строения морского ежа
Общая схема строения морского ежа
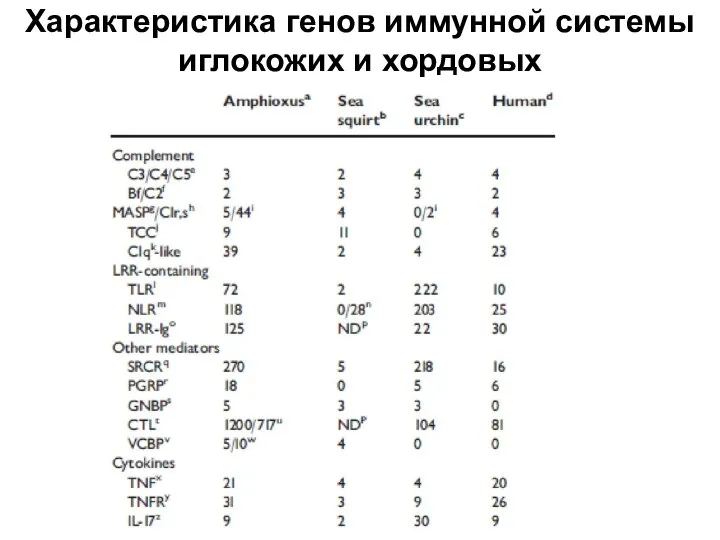
Слайд 33Характеристика генов иммунной системы иглокожих и хордовых
Характеристика генов иммунной системы иглокожих и хордовых
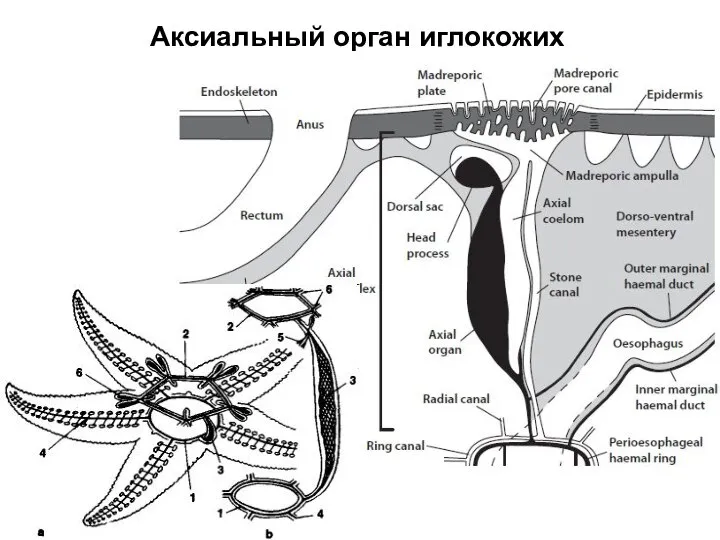
Слайд 34Аксиальный орган иглокожих
Аксиальный орган иглокожих
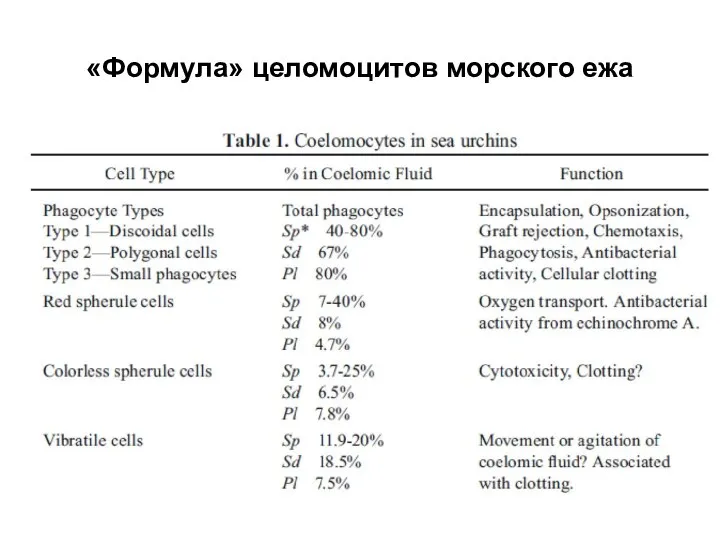
Слайд 35«Формула» целомоцитов морского ежа
«Формула» целомоцитов морского ежа
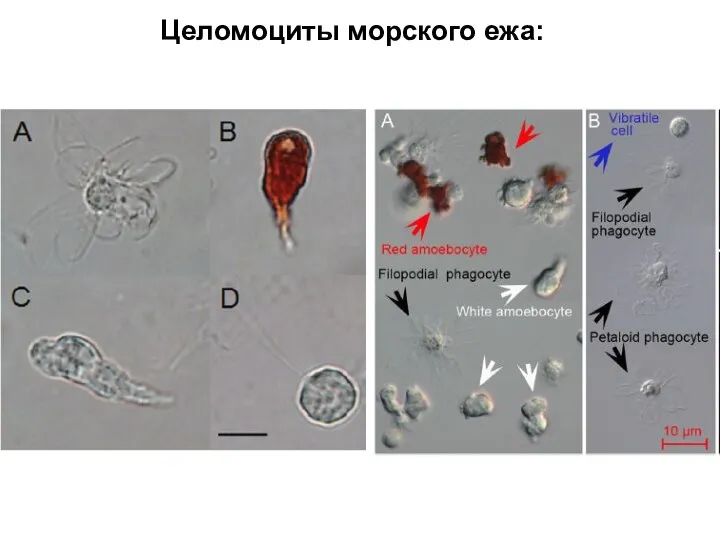
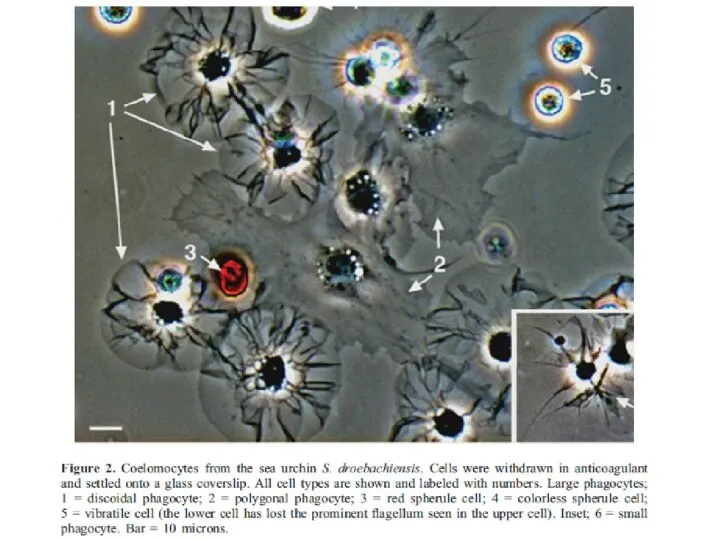
Слайд 36Целомоциты морского ежа:
Целомоциты морского ежа:
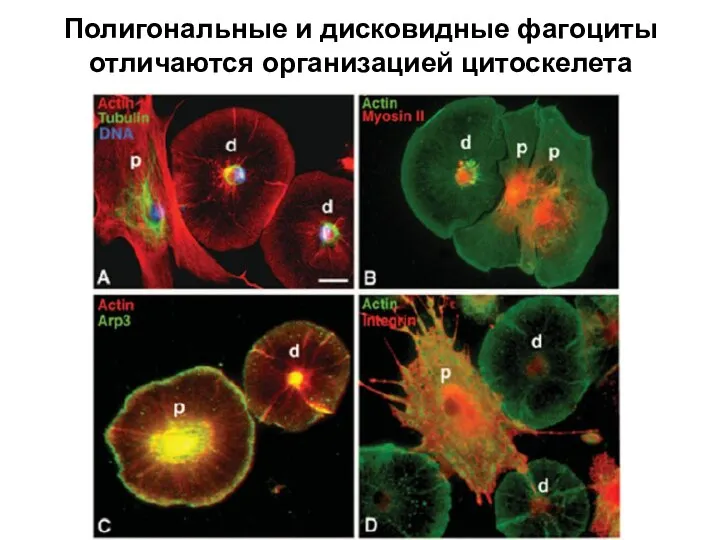
Слайд 38Полигональные и дисковидные фагоциты отличаются организацией цитоскелета
Полигональные и дисковидные фагоциты отличаются организацией цитоскелета
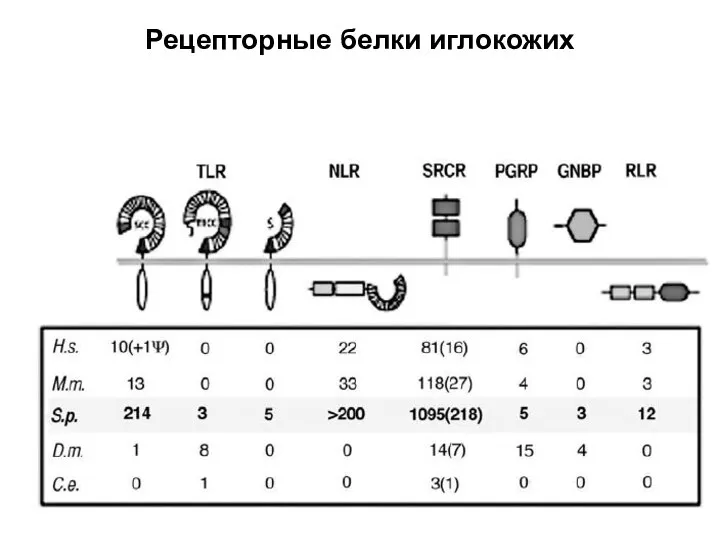
Слайд 39Рецепторные белки иглокожих
Рецепторные белки иглокожих
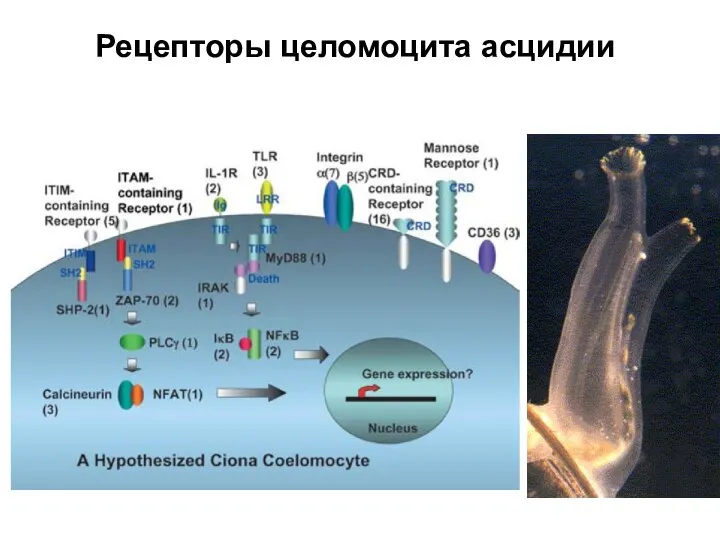
Слайд 40Рецепторы целомоцита асцидии
Рецепторы целомоцита асцидии
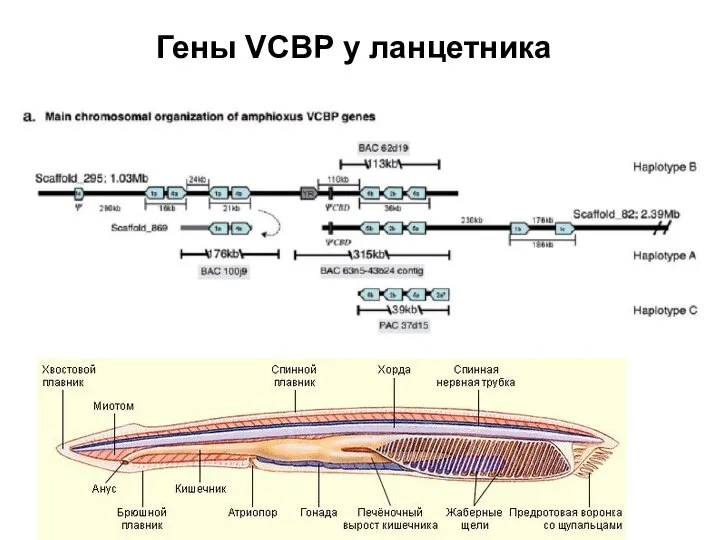
Слайд 41Гены VCBP у ланцетника
Гены VCBP у ланцетника
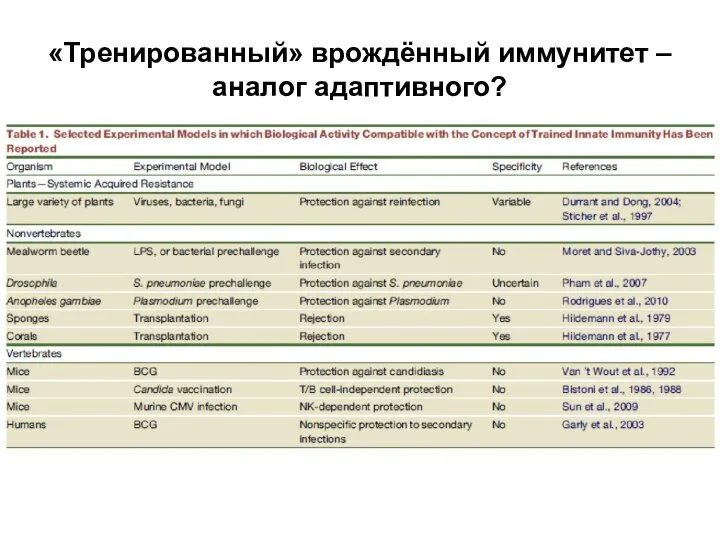
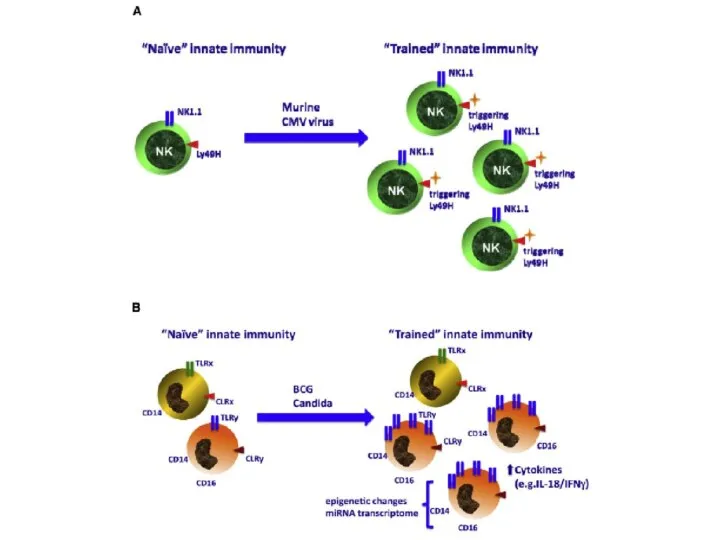
Слайд 42«Тренированный» врождённый иммунитет – аналог адаптивного?
«Тренированный» врождённый иммунитет – аналог адаптивного?
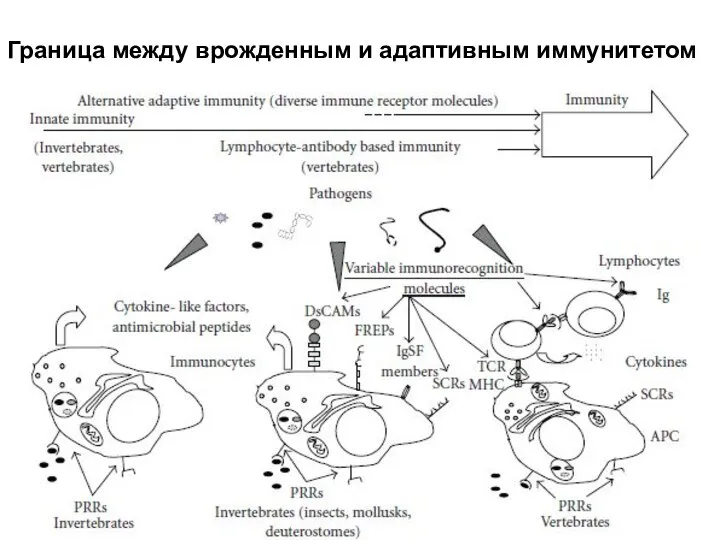
Слайд 44Граница между врожденным и адаптивным иммунитетом
Граница между врожденным и адаптивным иммунитетом
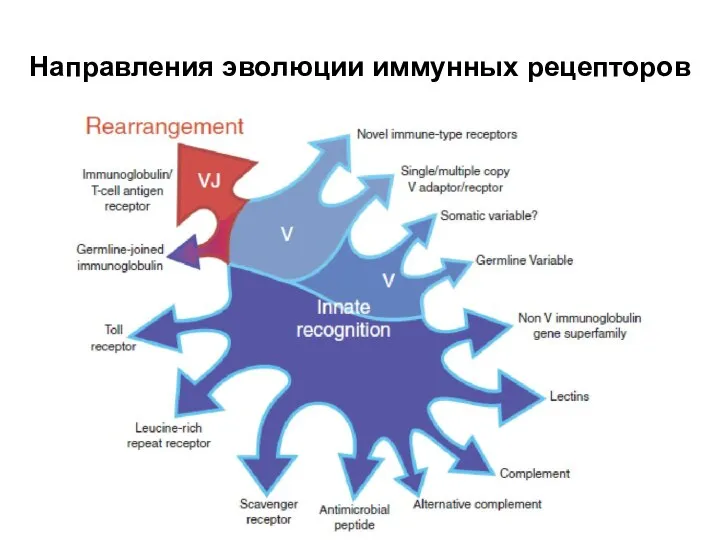
Слайд 45Направления эволюции иммунных рецепторов
Направления эволюции иммунных рецепторов
 Лептоспироз в Кировской области
Лептоспироз в Кировской области Зоб и тиреотоксикоз. Диффузно-токсический зоб
Зоб и тиреотоксикоз. Диффузно-токсический зоб Коленный сустав
Коленный сустав Оценка пациента и стартовая ИТ при политравме
Оценка пациента и стартовая ИТ при политравме Некроз ткани
Некроз ткани Аллергия к нежалящим насекомым
Аллергия к нежалящим насекомым УЗ - диагностика гнойно-воспалительных процессов в брюшной полости и мягких тканях
УЗ - диагностика гнойно-воспалительных процессов в брюшной полости и мягких тканях Экология микроорганизмов. (Лекция 2)
Экология микроорганизмов. (Лекция 2) Бронхиальная обструкция
Бронхиальная обструкция Антитела. Природа, виды, функции антител. Динамика продукции при первичном и вторичном иммунном ответе
Антитела. Природа, виды, функции антител. Динамика продукции при первичном и вторичном иммунном ответе Неврология в мире беременной
Неврология в мире беременной Хроническая сердечная недостаточность
Хроническая сердечная недостаточность Антидоты
Антидоты Полис. Компания Euromed Group - крупнейшая медицинская компания в Санкт-Петербурге
Полис. Компания Euromed Group - крупнейшая медицинская компания в Санкт-Петербурге Гемофилия қандай ауруға жатады
Гемофилия қандай ауруға жатады 病例展示
病例展示 Применение населением Могилевской области противомикробных лекарственных средств в домашних условиях
Применение населением Могилевской области противомикробных лекарственных средств в домашних условиях Оказание экстренной медицинской помощи пострадавшим с политравмой
Оказание экстренной медицинской помощи пострадавшим с политравмой Күйік және оның түрлері
Күйік және оның түрлері Эндодонтическое лечение
Эндодонтическое лечение Вегето-сосудистая дистония. Лекция 24
Вегето-сосудистая дистония. Лекция 24 Остеопороз эпидемиология, диагностика, лечение и профилактика
Остеопороз эпидемиология, диагностика, лечение и профилактика хрящевая ткань
хрящевая ткань Неоперативная хирургическая техника. Десмургия
Неоперативная хирургическая техника. Десмургия Кровотечение. Тест
Кровотечение. Тест Дүниежүзілік денсаулық сақтау ұйымының жаһандық мәселелерінің принциптері мен стратегиялары
Дүниежүзілік денсаулық сақтау ұйымының жаһандық мәселелерінің принциптері мен стратегиялары Врожденная краснуха
Врожденная краснуха СПИД. Пути передачи
СПИД. Пути передачи