- Надмолекулярные структуры

Содержание
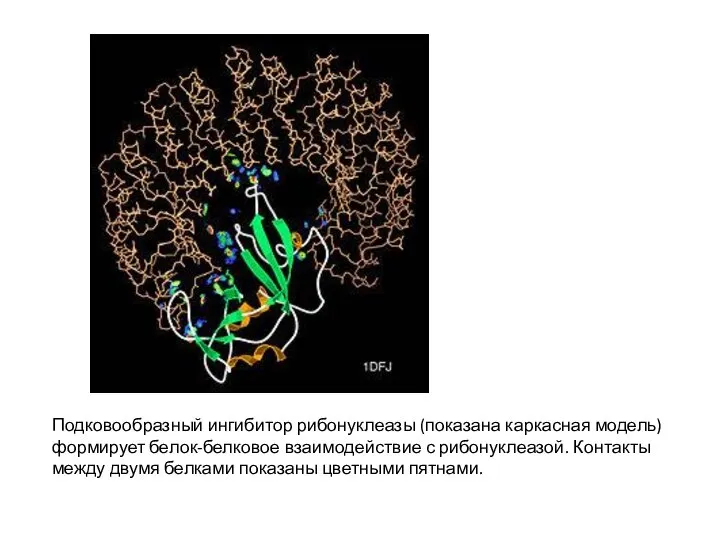
- 2. Подковообразный ингибитор рибонуклеазы (показана каркасная модель) формирует белок-белковое взаимодействие с рибонуклеазой. Контакты между двумя белками показаны
- 3. Примеры белок-белковых взаимодействий. Передача сигналов. Активность в клетке может регулироваться внеклеточными сигналами. Передача сигналов в клетке
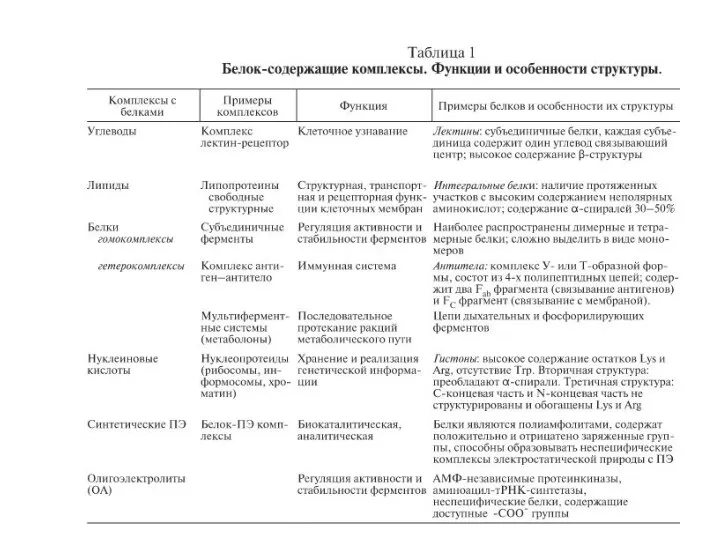
- 4. Типы белок-белковых взаимодействий. Комплексы белков могут образовывать гомо- и гетероолигомеры. Распространены комплексы фермент-ингибитор и антитело-антиген. Взаимодействия
- 5. Справка: Цитохром С - небольшой гем-содержащий белок, относится к классу цитохромов, содержит в структуре гем типа
- 6. Такие взаимодействия называют временными, например, некоторые белки с SH2-доменами пристыковываются к другим белкам, только когда их
- 7. Справка: убиквити́н (от англ. ubiquitous — вездесущий) — небольшой (8.5 кДа) консервативный белок эукариот, участвующий в
- 8. Справка: для многих белков, принимающих участие в передаче сигналов, характерно присутствие т.н. SH2 и SH3-доменов в
- 9. Особенности строения и функционирования олигомерных (т.е. состоят из нескольких пептидных цепей, называемых протомерами) белков на примере
- 10. Гемоглобин — олигомерный белок, функция которого регулируется различными лигандами. 1.Гемоглобин (НЬ) — сложный олигомерный белок, содержащийся
- 12. Комплексы белков с синтетическими полиэлектролитами (ПЭ) спонтанно образуются, главным образом, за счет электростатических взаимодействий. Белок–ПЭ комплексы
- 13. КОМПЛЕКСЫ БЕЛКОВ С ПРИРОДНЫМИ ОЛИГОАМИНАМИ. Многоточечное электростатическое взаимодействие с белком реализуется также при образовании комплексов с
- 15. Строение, свойства и функции клеточных мембран. В 1972 г. Сингер и Николсон (Singer, Nicolson) предложили жидкостно-мозаичную
- 16. На рисунке представлено плоскостное изображение жидкостно-мозаичной модели мембраны и ее трехмерная модель.
- 19. Надмолекулярные структуры внеклеточного матрикса. Значительную часть объема тканей занимает внеклеточное пространство, заполненное сложной сетью макромолекул, образующих
- 20. Протеогликаны – высокомолекулярные соединения, состоящие из белка (5–10%) и гликозаминогликанов (90–95%). Они образуют основное вещество межклеточного
- 21. В межклеточном матриксе присутствуют разные протеогликаны. Среди них есть очень крупные – например агрекан и ворсикан.
- 22. Липополисахарид (ЛПС; англ. lipopolysaccharide) — макромолекула, состоящая из полисахарида, ковалентно соединённого с липидом, основной компонент клеточной
- 23. Функция Липополисахарид (ЛПС, эндотоксин) — термостабильный компонент наружной части клеточной мембраны всех грамотрицательных микроорганизмов. ЛПС обеспечивает
- 24. Нуклеопротеиды — комплексы нуклеиновых кислот с белками. К нуклеопротеидам относятся устойчивые комплексы нуклеиновых кислот с белками,
- 25. Дезоксирибонуклеопротеиды. Хроматин — комплекс ДНК с гистонами в клетках эукариот. За счёт электростатического взаимодействия нить ДНК
- 26. Рибонуклеопротеиды. В клетках в наибольших количествах содержатся два класса рибонуклеопротеидов: Нуклеопротеидные комплексы рибосомальных РНК (рРНП) —
- 27. Справка: Сплайсосома — структура, состоящая из молекул РНК и белков и осуществляющая удаление некодирующих последовательностей (интронов)
- 28. Нуклеокапсиды вирусов. Нуклеокапсиды вирусов представляют собой достаточно плотно упакованные комплексы белков с нуклеиновой кислотой (ДНК или
- 30. Скачать презентацию
Слайд 2Подковообразный ингибитор рибонуклеазы (показана каркасная модель) формирует белок-белковое взаимодействие с рибонуклеазой. Контакты
Подковообразный ингибитор рибонуклеазы (показана каркасная модель) формирует белок-белковое взаимодействие с рибонуклеазой. Контакты
Слайд 3Примеры белок-белковых взаимодействий.
Передача сигналов. Активность в клетке может регулироваться внеклеточными сигналами. Передача
Примеры белок-белковых взаимодействий.
Передача сигналов. Активность в клетке может регулироваться внеклеточными сигналами. Передача

Слайд 4Типы белок-белковых взаимодействий.
Комплексы белков могут образовывать гомо- и гетероолигомеры. Распространены комплексы фермент-ингибитор
Типы белок-белковых взаимодействий.
Комплексы белков могут образовывать гомо- и гетероолигомеры. Распространены комплексы фермент-ингибитор

Слайд 5 Справка: Цитохром С - небольшой гем-содержащий белок, относится к классу цитохромов,
Справка: Цитохром С - небольшой гем-содержащий белок, относится к классу цитохромов,

Слайд 6Такие взаимодействия называют временными, например, некоторые белки с SH2-доменами пристыковываются к другим
Такие взаимодействия называют временными, например, некоторые белки с SH2-доменами пристыковываются к другим

Слайд 7Справка: убиквити́н (от англ. ubiquitous — вездесущий) — небольшой (8.5 кДа) консервативный белок эукариот, участвующий
Справка: убиквити́н (от англ. ubiquitous — вездесущий) — небольшой (8.5 кДа) консервативный белок эукариот, участвующий

Слайд 8Справка: для многих белков, принимающих участие в передаче сигналов, характерно присутствие т.н.
Справка: для многих белков, принимающих участие в передаче сигналов, характерно присутствие т.н.

Слайд 9Особенности строения и функционирования олигомерных (т.е. состоят из нескольких пептидных цепей, называемых
Особенности строения и функционирования олигомерных (т.е. состоят из нескольких пептидных цепей, называемых

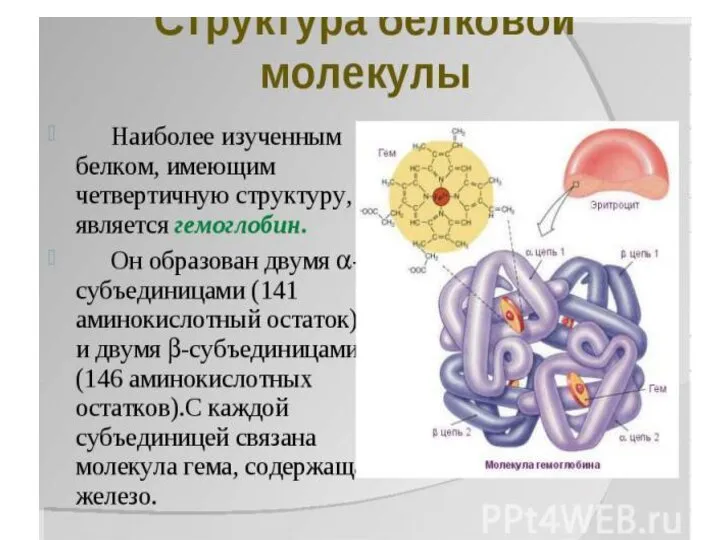
Слайд 10Гемоглобин — олигомерный белок, функция которого регулируется различными лигандами.
1.Гемоглобин (НЬ) — сложный
Гемоглобин — олигомерный белок, функция которого регулируется различными лигандами.
1.Гемоглобин (НЬ) — сложный

Слайд 12Комплексы белков с синтетическими полиэлектролитами (ПЭ) спонтанно образуются, главным образом, за счет
Комплексы белков с синтетическими полиэлектролитами (ПЭ) спонтанно образуются, главным образом, за счет

Слайд 13КОМПЛЕКСЫ БЕЛКОВ С ПРИРОДНЫМИ ОЛИГОАМИНАМИ.
Многоточечное электростатическое взаимодействие с белком реализуется также при
КОМПЛЕКСЫ БЕЛКОВ С ПРИРОДНЫМИ ОЛИГОАМИНАМИ.
Многоточечное электростатическое взаимодействие с белком реализуется также при

Слайд 15Строение, свойства и функции клеточных мембран.
В 1972 г. Сингер и Николсон (Singer,
Строение, свойства и функции клеточных мембран.
В 1972 г. Сингер и Николсон (Singer,

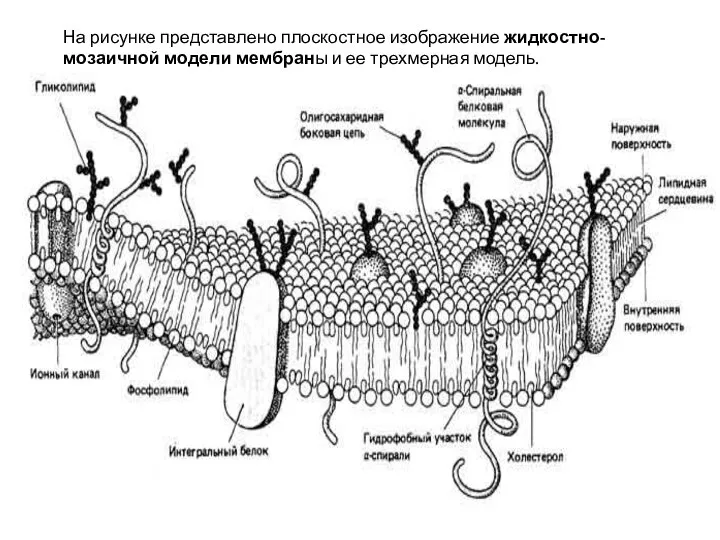
Слайд 16На рисунке представлено плоскостное изображение жидкостно-мозаичной модели мембраны и ее трехмерная модель.
На рисунке представлено плоскостное изображение жидкостно-мозаичной модели мембраны и ее трехмерная модель.
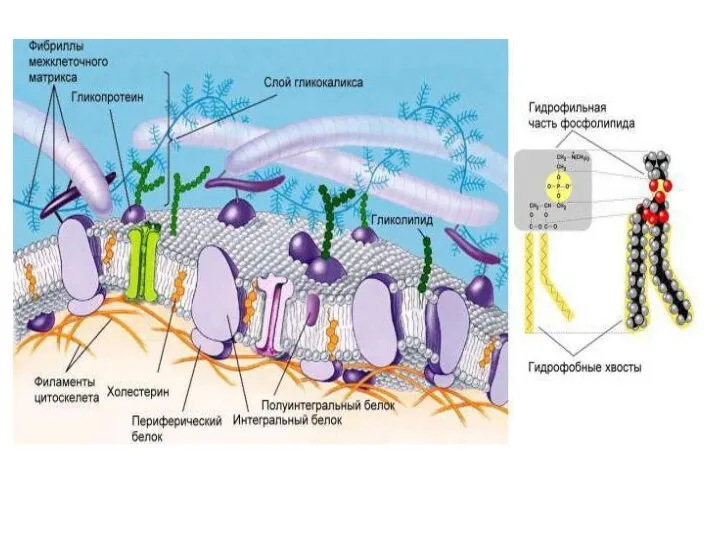

Слайд 19Надмолекулярные структуры внеклеточного матрикса.
Значительную часть объема тканей занимает внеклеточное пространство, заполненное
Надмолекулярные структуры внеклеточного матрикса.
Значительную часть объема тканей занимает внеклеточное пространство, заполненное

Слайд 20Протеогликаны – высокомолекулярные соединения, состоящие из белка (5–10%) и гликозаминогликанов (90–95%). Они
Протеогликаны – высокомолекулярные соединения, состоящие из белка (5–10%) и гликозаминогликанов (90–95%). Они

Слайд 21В межклеточном матриксе присутствуют разные протеогликаны. Среди них есть очень крупные –
В межклеточном матриксе присутствуют разные протеогликаны. Среди них есть очень крупные –

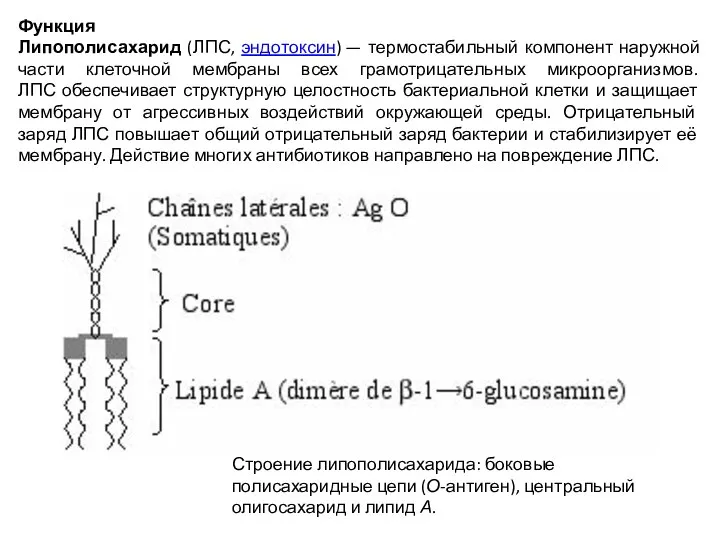
Слайд 22Липополисахарид (ЛПС; англ. lipopolysaccharide) — макромолекула, состоящая из полисахарида, ковалентно соединённого с липидом, основной
Липополисахарид (ЛПС; англ. lipopolysaccharide) — макромолекула, состоящая из полисахарида, ковалентно соединённого с липидом, основной

Слайд 23Функция
Липополисахарид (ЛПС, эндотоксин) — термостабильный компонент наружной части клеточной мембраны всех грамотрицательных микроорганизмов.
ЛПС
Функция
Липополисахарид (ЛПС, эндотоксин) — термостабильный компонент наружной части клеточной мембраны всех грамотрицательных микроорганизмов.
ЛПС
Слайд 24Нуклеопротеиды — комплексы нуклеиновых кислот с белками.
К нуклеопротеидам относятся устойчивые комплексы нуклеиновых кислот
Нуклеопротеиды — комплексы нуклеиновых кислот с белками.
К нуклеопротеидам относятся устойчивые комплексы нуклеиновых кислот

Слайд 25Дезоксирибонуклеопротеиды.
Хроматин — комплекс ДНК с гистонами в клетках эукариот. За счёт
электростатического взаимодействия
Дезоксирибонуклеопротеиды.
Хроматин — комплекс ДНК с гистонами в клетках эукариот. За счёт
электростатического взаимодействия

Слайд 26Рибонуклеопротеиды.
В клетках в наибольших количествах содержатся два класса рибонуклеопротеидов:
Нуклеопротеидные комплексы рибосомальных РНК
Рибонуклеопротеиды.
В клетках в наибольших количествах содержатся два класса рибонуклеопротеидов:
Нуклеопротеидные комплексы рибосомальных РНК

Слайд 27Справка: Сплайсосома — структура, состоящая из молекул РНК и белков и осуществляющая удаление
Справка: Сплайсосома — структура, состоящая из молекул РНК и белков и осуществляющая удаление

Слайд 28Нуклеокапсиды вирусов.
Нуклеокапсиды вирусов представляют собой достаточно плотно упакованные комплексы
белков с нуклеиновой кислотой
Нуклеокапсиды вирусов.
Нуклеокапсиды вирусов представляют собой достаточно плотно упакованные комплексы белков с нуклеиновой кислотой

 Раздел 1
Раздел 1 Карбоновые кислоты. Альдегиды
Карбоновые кислоты. Альдегиды Что объединяет вкус яблока, киви, садовой земляники?
Что объединяет вкус яблока, киви, садовой земляники? Минералы
Минералы Презентация на тему Увлекательные факты из жизни Менделеева
Презентация на тему Увлекательные факты из жизни Менделеева  Характеристика химического элемента по его положению в ПСХЭ Д.И.Менделева
Характеристика химического элемента по его положению в ПСХЭ Д.И.Менделева Чистые вещества и смеси. Классификация веществ. Оксиды
Чистые вещества и смеси. Классификация веществ. Оксиды Полиамиды (ПА)
Полиамиды (ПА) Презентация на тему Получение радиоактивных изотопов и их применение
Презентация на тему Получение радиоактивных изотопов и их применение  Кислород. Общая характеристика подгруппы на примере кислорода и серы
Кислород. Общая характеристика подгруппы на примере кислорода и серы Обобщение и повторение за курс основной школы по химии. 9 класс
Обобщение и повторение за курс основной школы по химии. 9 класс хімія
хімія Химическое равновесие
Химическое равновесие Основы химической термодинамики. Основные понятия химической термодинамики. Лекция 1
Основы химической термодинамики. Основные понятия химической термодинамики. Лекция 1 Карбон
Карбон Биологическая роль III(A) группы
Биологическая роль III(A) группы Презентация на тему Углекислый газ
Презентация на тему Углекислый газ  Железо и его сплавы
Железо и его сплавы Пиримидин (шестичленный гетероцикл с двумя атомами азота)
Пиримидин (шестичленный гетероцикл с двумя атомами азота) Алкины
Алкины Природные смолы: канифоль, янтарь, сандарак (2)
Природные смолы: канифоль, янтарь, сандарак (2) Валентность химических элементов
Валентность химических элементов Характеристика 1-й аналитической группы анионов
Характеристика 1-й аналитической группы анионов Влияние инициаторов на реакцию полимеризации стирола
Влияние инициаторов на реакцию полимеризации стирола Основной государственный экзамен. Химия 2022. Задание 2
Основной государственный экзамен. Химия 2022. Задание 2 Алкины. Способы получения
Алкины. Способы получения Типы химических реакций
Типы химических реакций Минералы алюминия
Минералы алюминия