- romanchuk

Содержание
- 2. СОДЕРЖАНИЕ Ведение 1. Общая характеристика белковых молекул 1.1. Элементный состав белков 1.2. Аминокислотный состав белков 1.2.1.
- 3. 3.Физико-химические свойства белков 3.1. Общие свойства 3.2. Электрические свойства белковых молекул 3.3. Растворимость белков 3.4. Денатурация
- 4. Условные обозначения Глицин Гиперссылка Глоссарий Шаг назад Шаг вперед К содержанию Видеофрагмент - - - -
- 5. Ведение Белки или протеины (от греч. protos–первый, важнейший) – это высокомолекулярные азотсодержащие органические вещества, линейные гетерополимеры,
- 6. Кроме понятия «белок», в химии встречается термины «пептид» и «полипептид». Пептидом обычно называют олигомер, состоящий не
- 7. Совокупность белковых молекул имеющихся в организме называется протеомом. Термин "протеом" предложил в 1994 году австралийский исследователь
- 8. Основные химические элементы, входящие в состав белков С(углерод) – 50-55% O(кислород) – 21-23% H(водород) – 6,5-7,3%
- 9. 1.2. Аминокислотный состав белков Белки построены в основном двадцатью стандартными (основными, природными, белковыми, кодируемыми) α -
- 10. Аминокислоты входящие в состав белка являются α-аминокислотами и представляют собой производные карбоновых кислот, у которых один
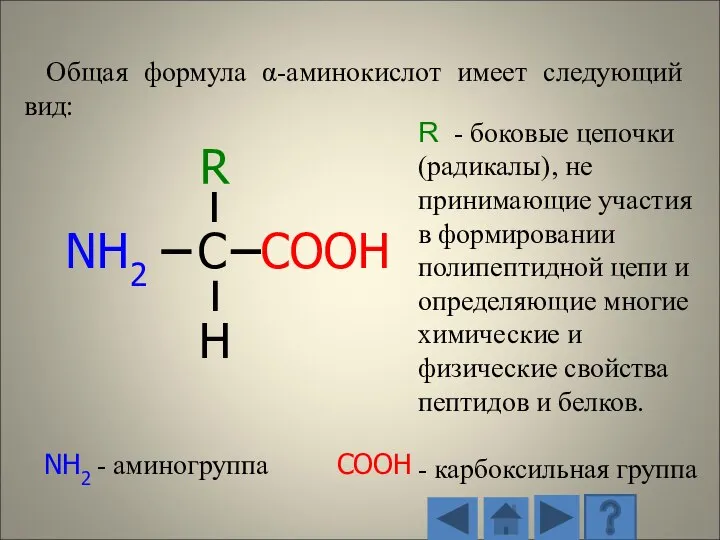
- 11. Общая формула α-аминокислот имеет следующий вид: R - боковые цепочки (радикалы), не принимающие участия в формировании
- 12. Природные аминокислоты имеют тривиальные названия, которые происходят иногда от источника из которого аминокислота была выделена. Например,
- 13. 1.2.1. Характеристика стандартных аминокислот Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H
- 14. Структурная формула Пространственное строение радикала (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп,
- 15. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 16. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 17. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 18. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 19. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 20. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 21. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 22. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 23. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 24. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 25. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 26. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 27. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 28. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 29. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 30. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 31. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 32. Структурная формула Пространственное строение (Серыми кружками показаны атомы С, светло-серыми — H атомы боковых групп, голубыми
- 33. 20 α-аминокислот Глицин (G) Глютаминовая кислота (E) Аспарагиновая кислота (D) Метионин (M) Треонин (T) Серин (S)
- 34. Важным свойством аминокислот является оптическая изомерия. Это свойство связано с наличием в молекуле всех природных аминокислот,
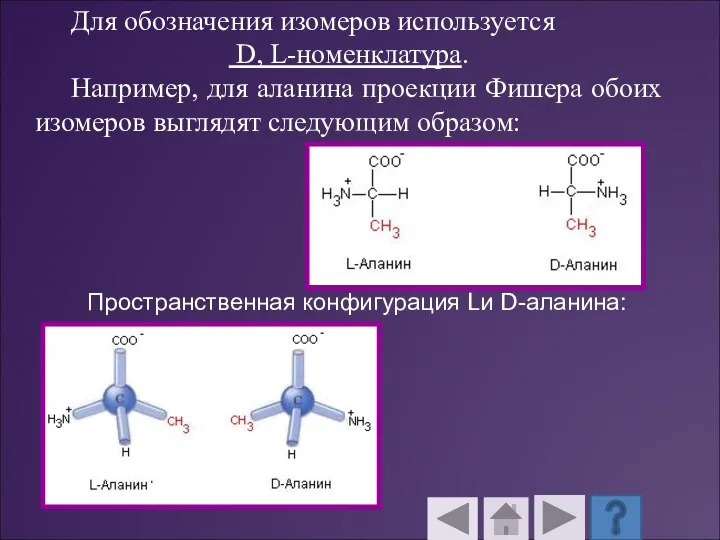
- 35. Для обозначения изомеров используется D, L-номенклатура. Например, для аланина проекции Фишера обоих изомеров выглядят следующим образом:
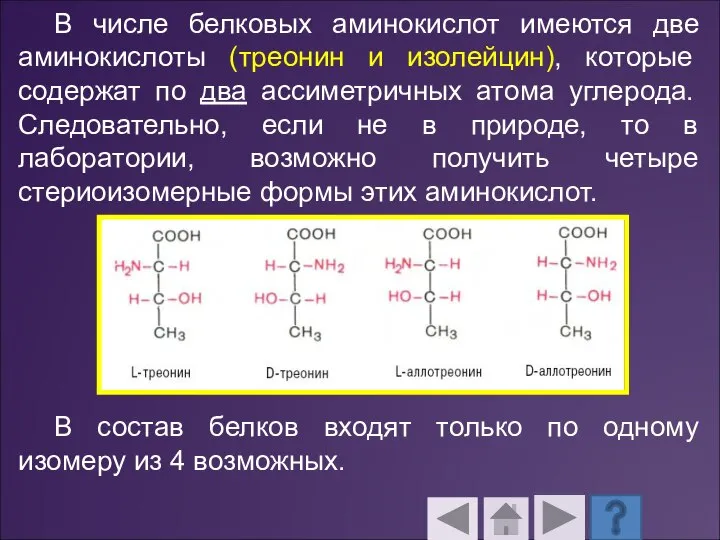
- 36. В числе белковых аминокислот имеются две аминокислоты (треонин и изолейцин), которые содержат по два ассиметричных атома
- 37. L - и D-изомеры могут взаимно превращаться друг в друга. Этот процесс называется рацемизацией. Все аминокислоты,
- 38. В белке зубов – дентине – L-аспарагиновая кислота самопроизвольно рацемизуется при температуре человеческого тела со скорость
- 39. 1.2.3. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА АМИНОКИСЛОТ Физические свойства Аминокислоты представляют собой кристаллические вещества с высокими (выше 250С0) температурами
- 40. Химические свойства Аминокислоты содержат амино- и карбоксильную группы. В силу этого они проявляют амфотерные свойства, то
- 41. Химические свойства При растворении аминокислоты в воде, например, глицина, его карбоксильная группа диссоциирует с образованием иона
- 42. Кислотно-основные свойства не ионная форма; идеализированная аминокислота цвиттер-ион
- 43. Кислотно-основные свойства В кислой среде аминокислота, присоединяя ион водорода, образует катион: В щелочной среде образуется анион:
- 44. Важной особенностью аминокислот является их способность к поликонденсации, приводящей к образованию полиамидов, в том числе пептидов
- 45. Для α-аминокислот характерны следующие химические реакции: реакции по карбоксильной группе; реакции по аминогруппе; биологически важные реакции.
- 46. 1.2.4.Классификация α-аминокислот В зависимости от подхода можно выделить следующие типы классификаций аминокислот:
- 47. Химическая классификация В соответствии с химической структурой радикала аминокислоты делят на алифатические, ароматические, а также аминокислоты,
- 48. Алифатические Моноаминомонокарбоновые: аланин, валин, глицин, изолейцин, лейцин. Оксимоноаминокарбоновые: серин, треонин. Моноаминодикарбоновые: аспарагиновая кислота, глутаминовая кислота, за
- 49. Ароматические: фенилаланин, тирозин - Гетероциклические: триптофан, гистидин, пролин (также входит в группу иминокислот) - Иминокислоты: пролин
- 50. Физико-химическая классификация основана на полярности радикалов (R-групп), т.е. способности их к взаимодействию с водой при физиологических
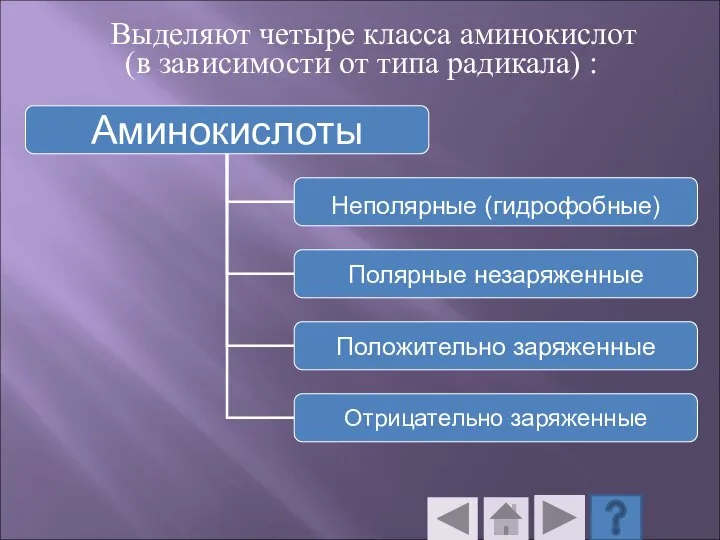
- 51. Выделяют четыре класса аминокислот (в зависимости от типа радикала) :
- 52. К неполярным (гидрофобным) относятся аминокислоты с неполярными алифатическими (аланин, валин, лейцин, изолейцин) или ароматическими (фенилаланин и
- 53. Полярные незаряженные аминокислоты в сравнении с неполярными лучше растворяются в воде, более гидрофильны, так как их
- 54. К положительно заряженным аминокислотам принадлежат лизин, гистидин и аргинин, в ионизированном виде они имеют суммарный положительный
- 55. К отрицательно заряженным аминокислотам относятся аспарагиновая и глутаминовая кислоты. Они содержат по две карбоксильные и по
- 56. Биологическая классификация основана на способности аминокислот, принимающие участие в построении белков человеческого тела, синтезироваться в нашем
- 57. Заменимые аминокислоты могут синтезироваться в организме человека (их 10). Незаменимые аминокислоты, их еще называют "эссенциальные» не
- 59. Незаменимые аминокислоты входят часто в состав пищевых добавок, используются в качестве лекарственных препаратов.
- 60. C 1981 г. в США разрешено использование низкокалорийной добавки для придания продуктам сладкого вкуса - аспартама.
- 61. Аргинин содержится в твороге, морепродуктах,грецких орехах,шоколаде. Валин содержится в зерновых, мясе, грибах, арахисе, сое Гистидин содержится
- 62. Интересно знать! Исключительно важную роль играет сбалансированность питания человека по аминокислотам. При недостатке незаменимых аминокислот в
- 63. 1.2.5. Производные аминокислот Нестанда́ртные аминокисло́ты — аминокислоты, которые, хотя и не входят в список встречающихся во
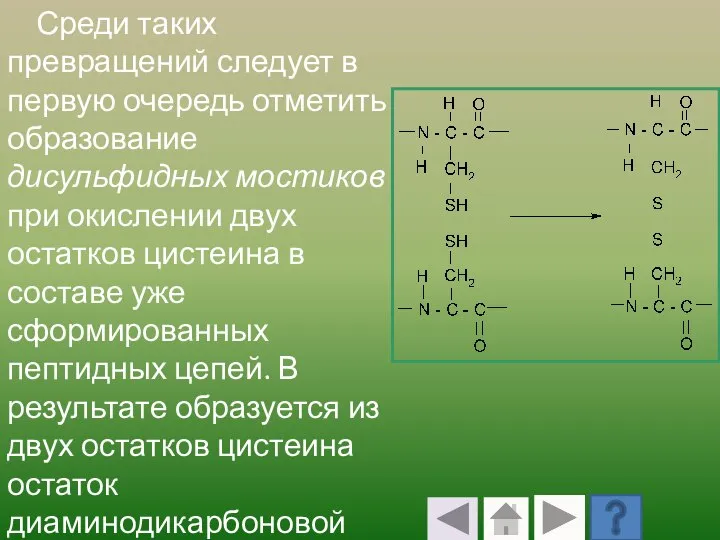
- 64. Среди таких превращений следует в первую очередь отметить образование дисульфидных мостиков при окислении двух остатков цистеина
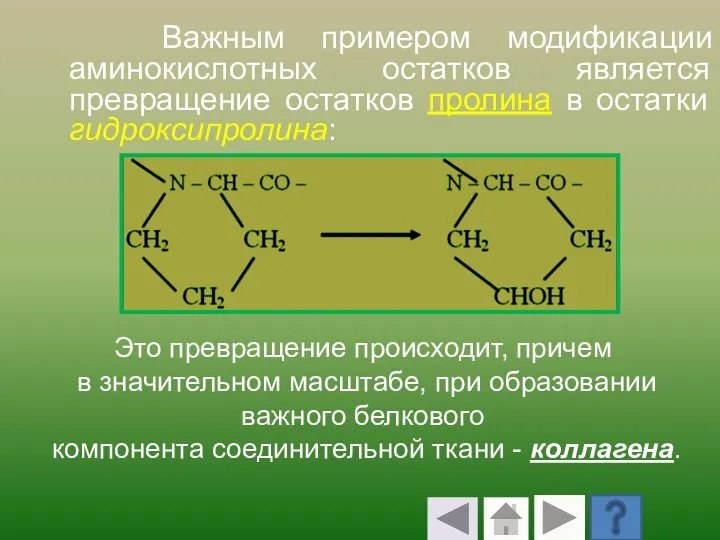
- 65. Важным примером модификации аминокислотных остатков является превращение остатков пролина в остатки гидроксипролина: Это превращение происходит, причем
- 66. Еще одним весьма важным видом модификации белков является фосфорилирование гидроксогрупп остатков серина, треонина и тирозина, например:
- 67. По составу белковой молекулы 1.3. Простые и сложные белки
- 68. Простые белки Простые белки состоят только из аминокислотных остатков и не содержат других химических составляющих. Альбумины
- 69. Альбумины (от лат. albumen, род. падеж albuminis - белок) - водорастворимые глобулярные белки, входящие в состав
- 70. Сывороточный альбумин составляет примерно 60% от общего количества белков плазмы крови. Молекулярный вес его - 69
- 71. Глобулины — слаборастворимые в воде белки, растворимы в разбавленных растворах солей, входят в состав растительных и
- 72. Гистоны белки, обладающие щелочными свойствами и входящие в состав комплексов с ДНК (хроматин, нуклеосомы) в ядрах
- 73. Сложные белки содержат небелковый компонент — простетическую группу. В зависимости от химической природы простетической группы протеиды
- 74. Металлопротеины белки, в составе которых имеются ионы металлов (медь, железо, цинк, молибден, марганец и др.). Строение
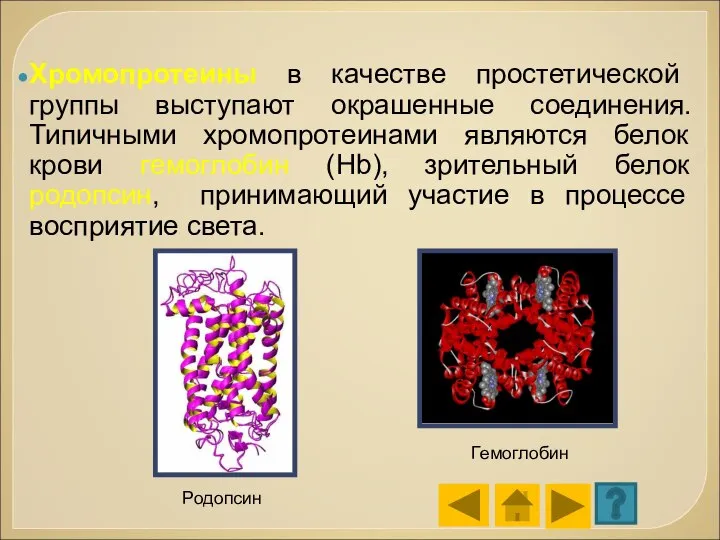
- 75. Хромопротеины в качестве простетической группы выступают окрашенные соединения. Типичными хромопротеинами являются белок крови гемоглобин (Hb), зрительный
- 76. Фосфопротены в своем составе содержат остатки фосфорной кислоты, связанные с гидроксильной группой аминокислотных остатков сложноэфирной связью.
- 77. Фосфопротены могут терять фосфатную группу (дефосфорилирование), могут при определенных условиях быть снова фосфорилированы. Одни белки проявляют
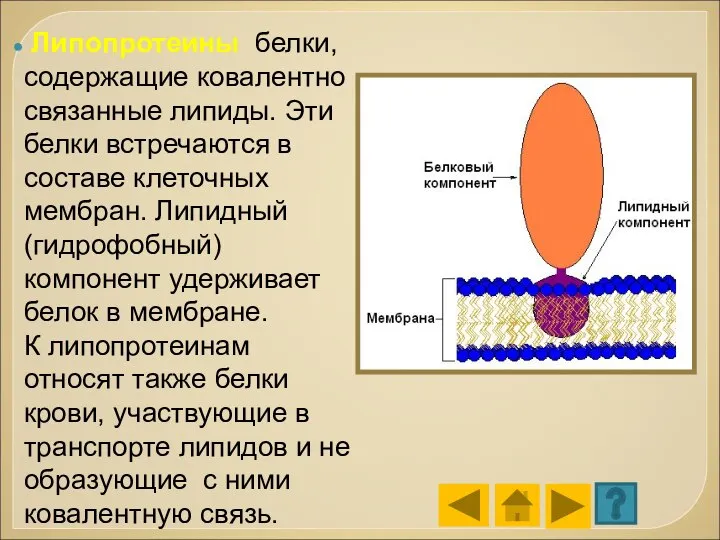
- 78. Липопротеины белки, содержащие ковалентно связанные липиды. Эти белки встречаются в составе клеточных мембран. Липидный (гидрофобный) компонент
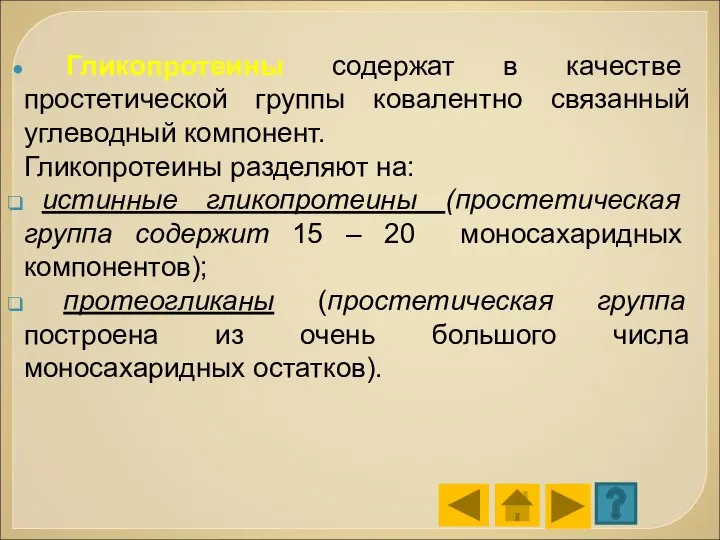
- 79. Гликопротеины содержат в качестве простетической группы ковалентно связанный углеводный компонент. Гликопротеины разделяют на: истинные гликопротеины (простетическая
- 81. Гликопротеины широко распространены в природе. Они входят в состав клеточных мембран, клеточных стенок, соединительной ткани, встречаются
- 82. Нуклеопротеины - состоят из белков и нуклеиновых кислот, крайние рассматриваются как простетические группы. ДНП-дезоксирибонуклеопротеиды. РНП-рибонуклеопротеиды. ДНП
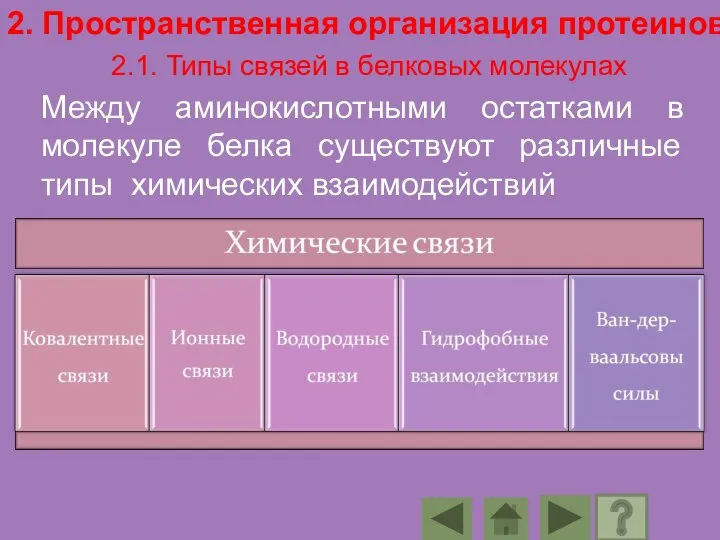
- 83. 2. Пространственная организация протеинов 2.1. Типы связей в белковых молекулах Между аминокислотными остатками в молекуле белка
- 84. Ковалентные связи
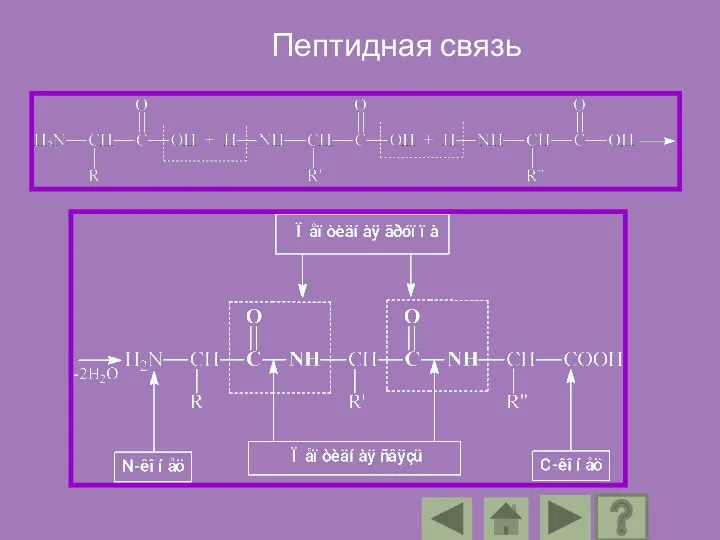
- 85. По своей природе пептидная связь является ковалентной. Ее образование происходит за счет α-аминогруппы одной аминокислоты и
- 86. Название пептида образуется из названия аминокислотных остатков пептидной цепи путем замены их окончаний на окончание –ил
- 87. Пептидная связь
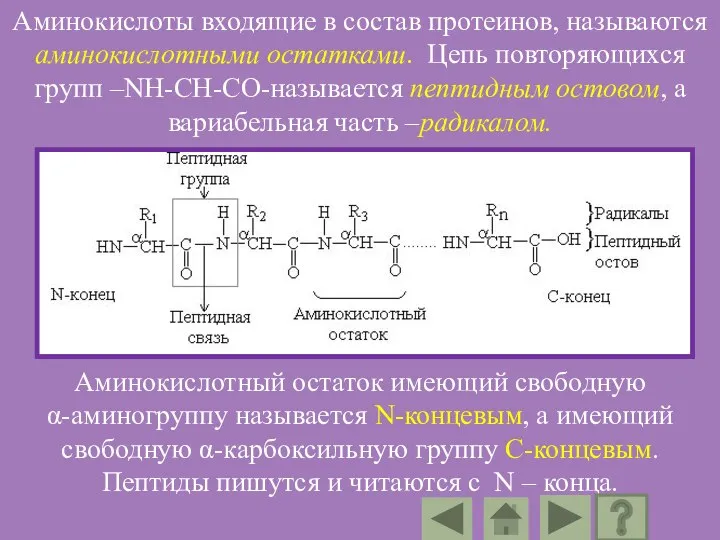
- 88. Аминокислоты входящие в состав протеинов, называются аминокислотными остатками. Цепь повторяющихся групп –NH-CH-CO-называется пептидным остовом, а вариабельная
- 89. В пептидах выделяют особую структуру – пептидную группу. Ее образуют атомы кислорода, углерода, азота и водорода.
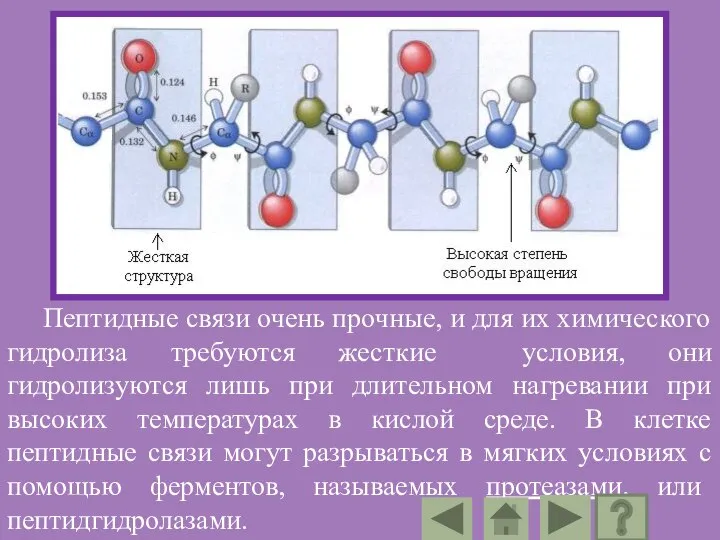
- 90. Свойства пептидной группы: Пептидная группа имеет жесткую планарную структуру, т.е. все атомы, входящие в нее, располагаются
- 91. Пептидные связи очень прочные, и для их химического гидролиза требуются жесткие условия, они гидролизуются лишь при
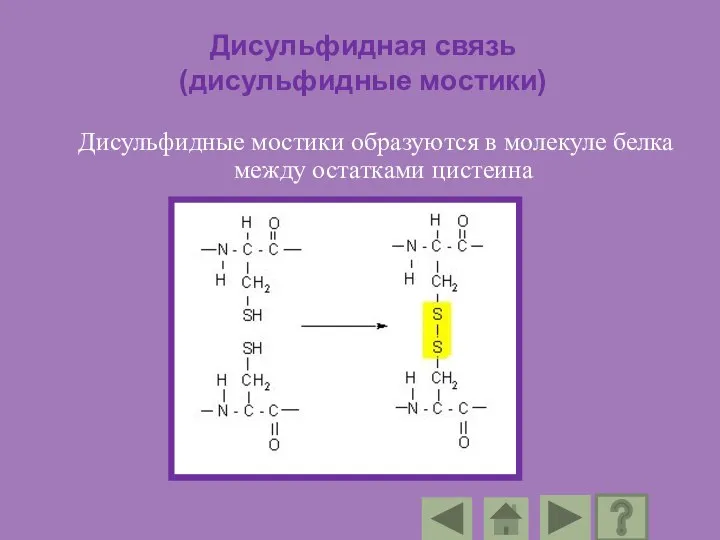
- 92. Дисульфидная связь (дисульфидные мостики) Дисульфидные мостики образуются в молекуле белка между остатками цистеина
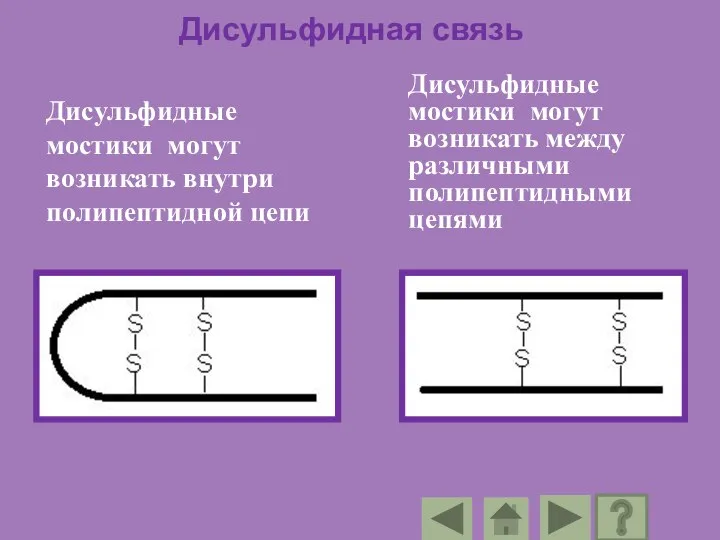
- 93. Дисульфидная связь Дисульфидные мостики могут возникать внутри полипептидной цепи Дисульфидные мостики могут возникать между различными полипептидными
- 94. Интересно знать! С помощью химической завивки волосам можно придать другую форму. Для этого волосы обрабатывают раствором
- 95. Ионные связи Ионные связи возникают между радикалами аминокислотных остатков, имеющих противоположные заряды, например, между положительно заряженной
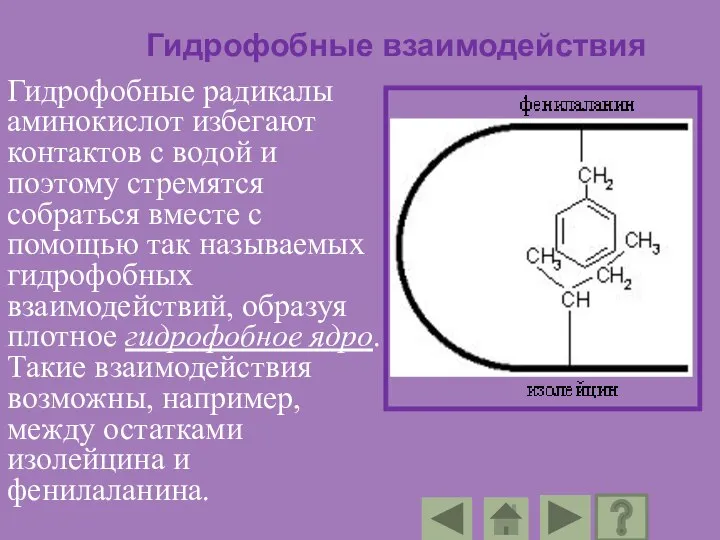
- 96. Гидрофобные взаимодействия Гидрофобные радикалы аминокислот избегают контактов с водой и поэтому стремятся собраться вместе с помощью
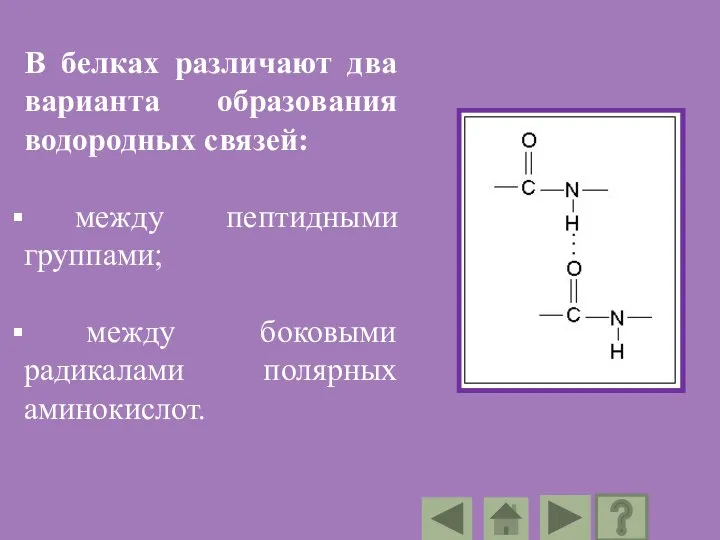
- 97. Водородные связи Водородная связь в молекуле белка образуется между имеющим частично положительный заряд атомом водорода одной
- 98. В белках различают два варианта образования водородных связей: между пептидными группами; между боковыми радикалами полярных аминокислот.
- 99. Ван-дер-ваальсовы силы Ван-дер-ваальсовы силы имеют электростатическую природу. Они возникают между разноименными полюсами диполя. В молекуле белка
- 100. Рассмотренные выше химические связи принимают участие в формировании структуры белковых молекул
- 101. 2.2. Уровни организации белковых молекул
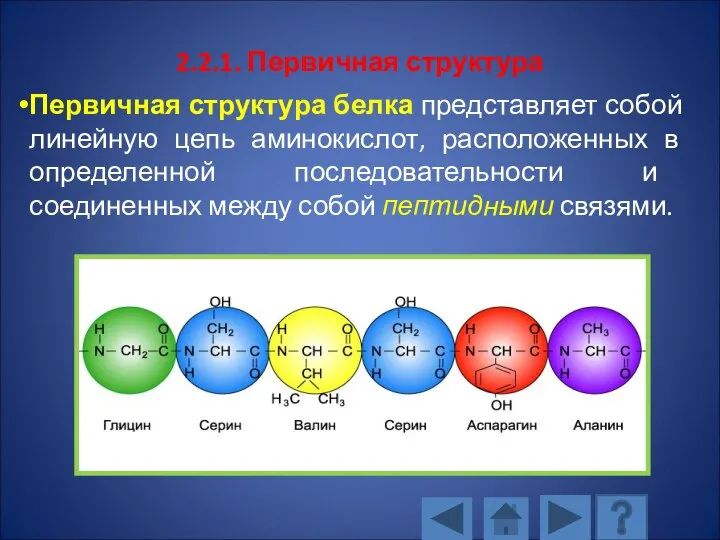
- 102. 2.2.1. Первичная структура Первичная структура белка представляет собой линейную цепь аминокислот, расположенных в определенной последовательности и
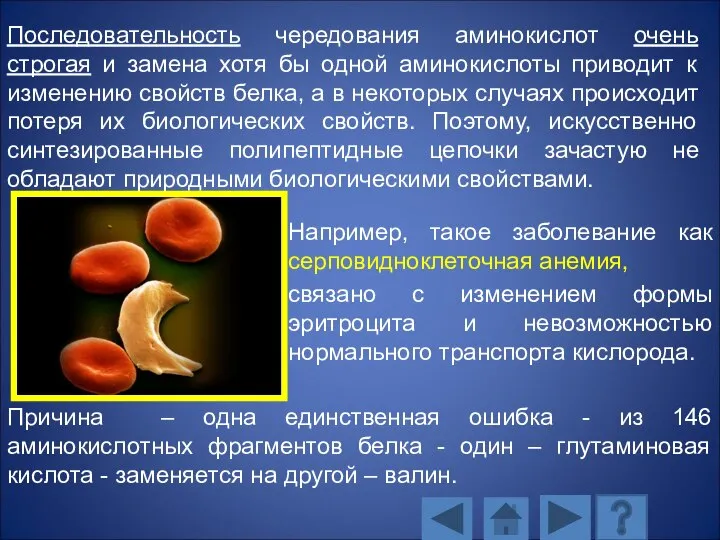
- 103. Первичная структура белка характеризуется: аминокислотным составом; последовательностью аминокислотных остатков; количеством аминокислотных остатков.
- 104. Например, такое заболевание как серповидноклеточная анемия, связано с изменением формы эритроцита и невозможностью нормального транспорта кислорода.
- 105. Фред Сенгер Фред Сенгер определил аминокислотную последовательность инсулина, продемонстрировав таким способом, что белки — это линейные
- 106. 2.2.2. Вторичная структура Вторичная структура белка образуется в процессе укладки первичной структуры в пространстве в виде
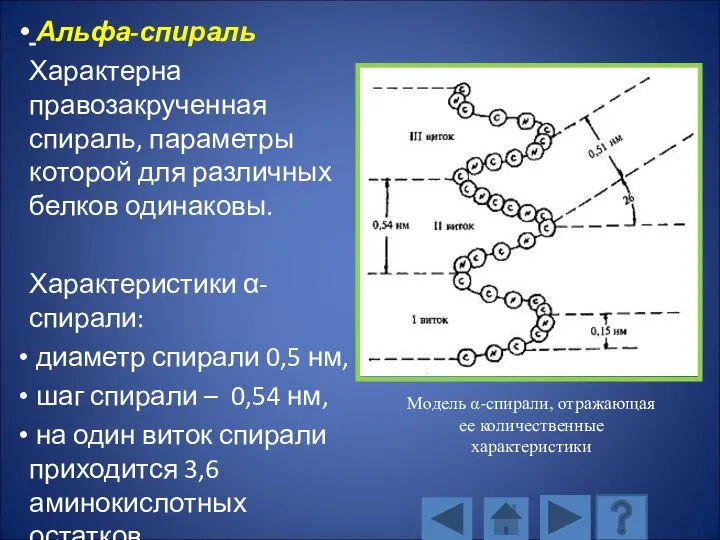
- 107. Альфа-спираль Характерна правозакрученная спираль, параметры которой для различных белков одинаковы. Характеристики α-спирали: диаметр спирали 0,5 нм,
- 108. Альфа-спираль Водородные связи, фиксирующие альфа-спираль расположены параллельно оси спирали и многократно повторяются, поэтому прочно удерживают спиралеобразную
- 109. Некоторые аминокислотные остатки препятствуют образованию α-спирали: Например, если в цепи подряд расположено несколько положительно или отрицательно
- 110. В остатке пролина при атоме азота, образующем пептидную связь с другой аминокислотой, нет атома водорода, поэтому
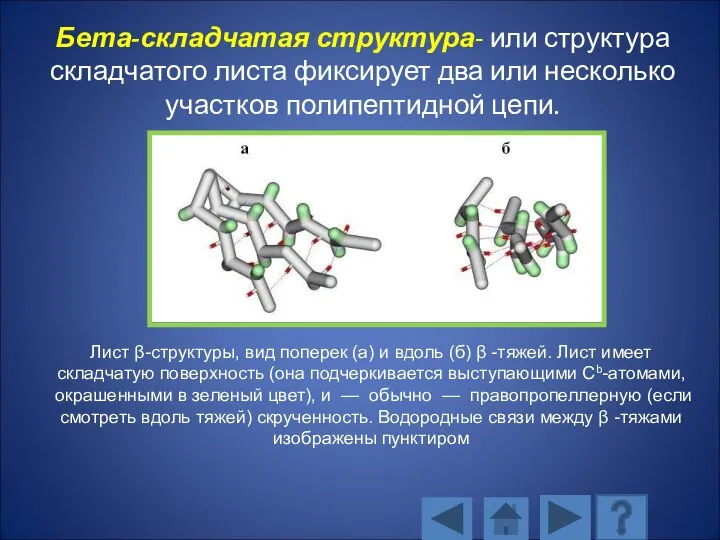
- 111. Бета-складчатая структура- или структура складчатого листа фиксирует два или несколько участков полипептидной цепи. Лист β-структуры, вид
- 112. Цепи могут быть параллельны или антипараллельны.
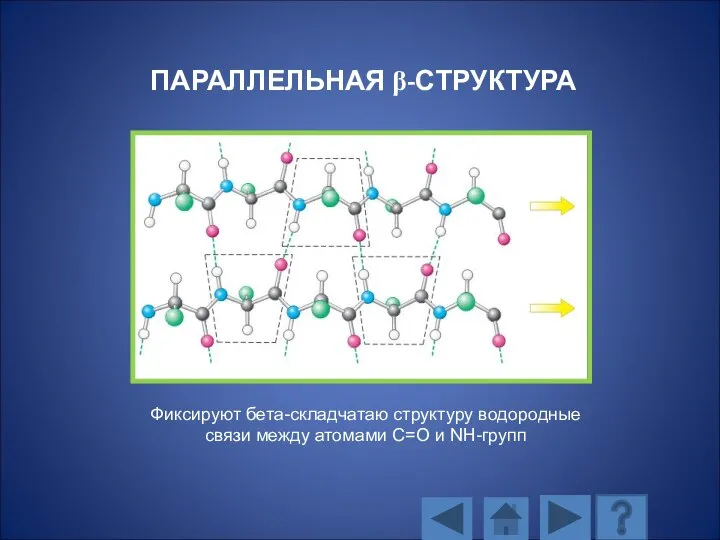
- 113. ПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА Фиксируют бета-складчатаю структуру водородные связи между атомами С=О и NH-групп
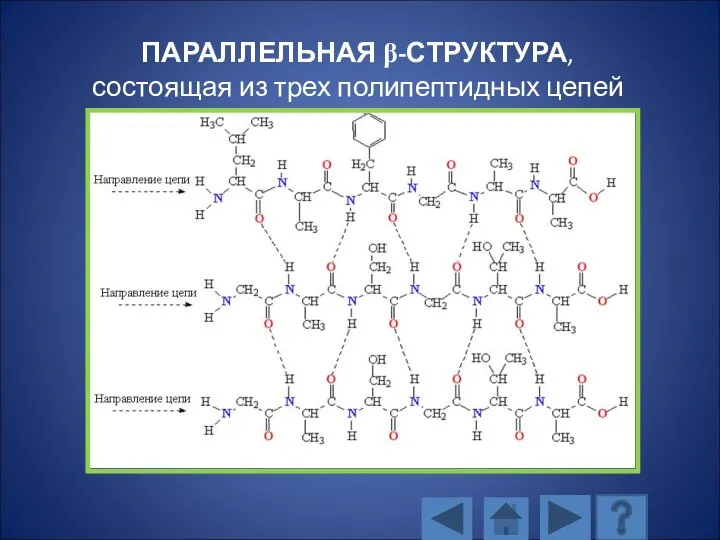
- 114. ПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА, состоящая из трех полипептидных цепей
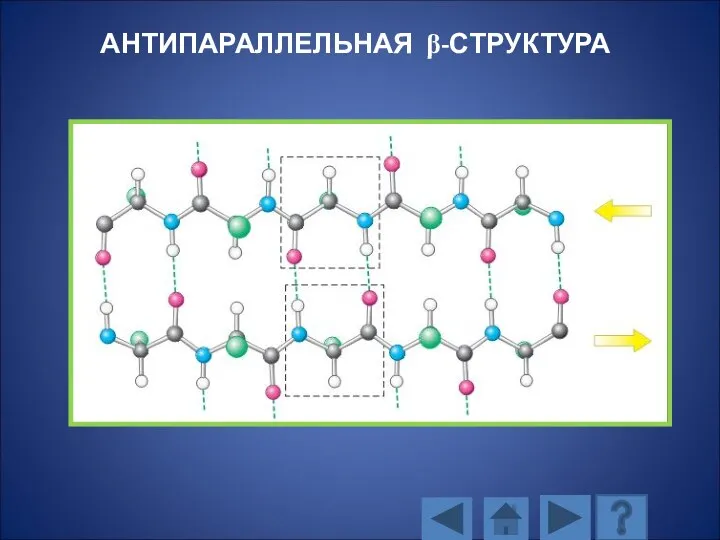
- 115. АНТИПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА
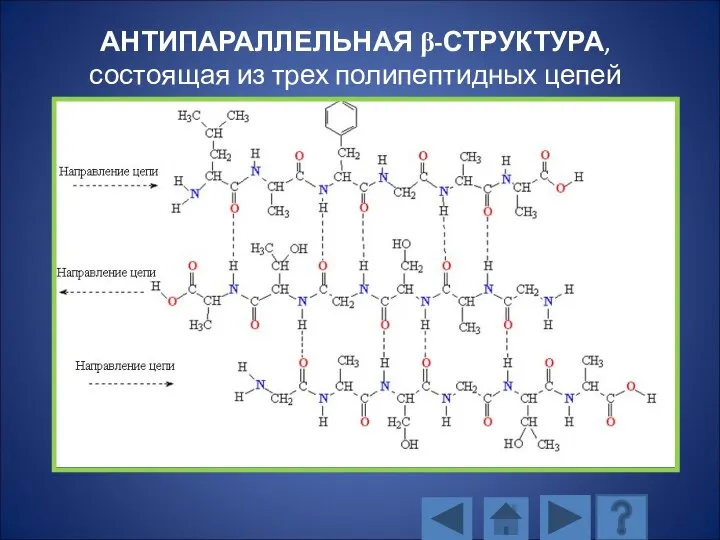
- 116. АНТИПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА, состоящая из трех полипептидных цепей
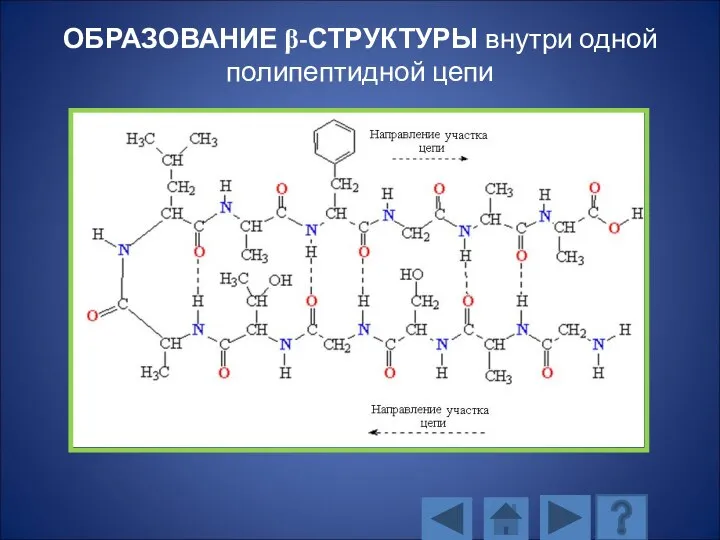
- 117. ОБРАЗОВАНИЕ β-СТРУКТУРЫ внутри одной полипептидной цепи
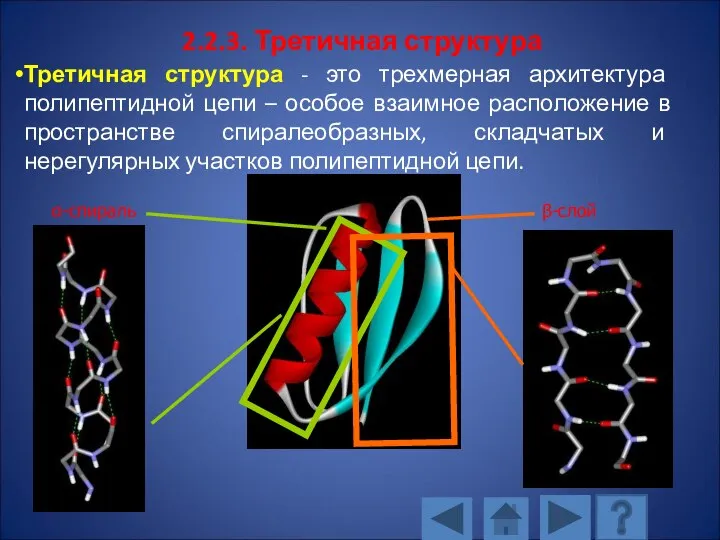
- 118. 2.2.3. Третичная структура Третичная структура - это трехмерная архитектура полипептидной цепи – особое взаимное расположение в
- 119. У разных белков третичной структуры различна. В формировании третичной структуры участвуют дисульфидные связи и все слабые
- 120. В фибриллярных белках (например, коллаген, эластин) молекулы имеют вытянутую форму и обычно формируют волокнистые структуры тканей.
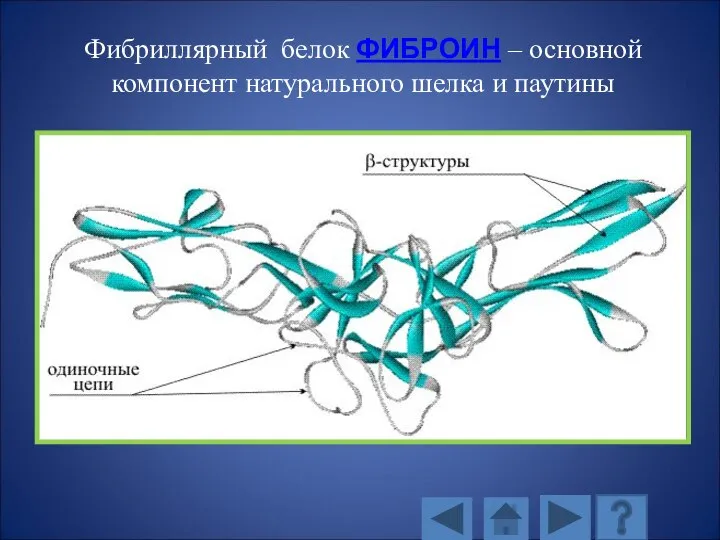
- 121. Фибриллярный белок ФИБРОИН – основной компонент натурального шелка и паутины
- 122. В организме млекопитающих коллаген —составляет 25% общего белка. Этот белок имеет необычный аминокислотный состав: -1/З составляет
- 123. В глобулярных белках молекулы имеют форму шара или эллипса. Обычно гидрофобные участки находятся в глубине молекулы,
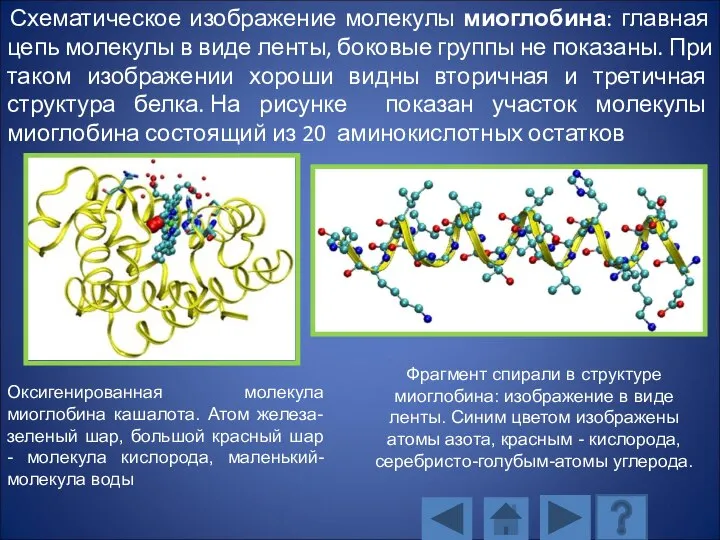
- 124. Миоглобин – кислород-связывающий белок. Молекула состоит из одной полипептидной цепи, состоящей из 153 аминокислотных остатков, и
- 125. Схематическое изображение молекулы миоглобина: главная цепь молекулы в виде ленты, боковые группы не показаны. При таком
- 126. Можем ли мы определить трехмерную форму белка по его аминокислотной последовательности? Сейчас это пока невозможно. Предопределение
- 127. 2.2.4. Четвертичная структура Четвертичная структура - субъединичная структура белка, представляющая взаимное расположение нескольких полипептидных цепей (протомеров)
- 128. Число протомеров может варьировать в широких пределах. Чаще в молекуле олигомерного белка содержится 2 или 4
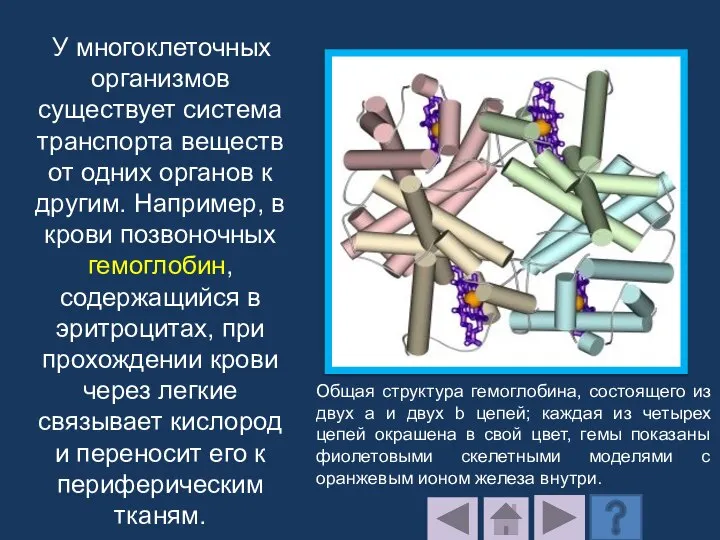
- 129. Молекула гемоглобина состоит из гема и четырех полипептидных цепей двух различных типов – двух α-цепей и
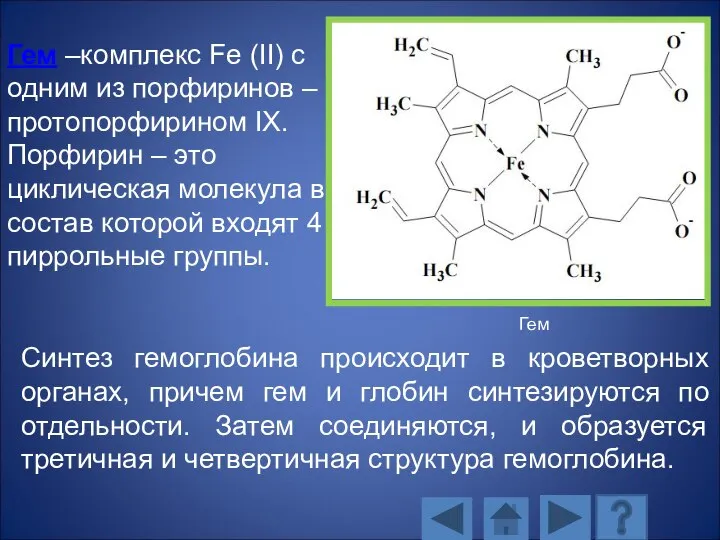
- 130. Гем –комплекс Fe (II) с одним из порфиринов – протопорфирином IX. Порфирин – это циклическая молекула
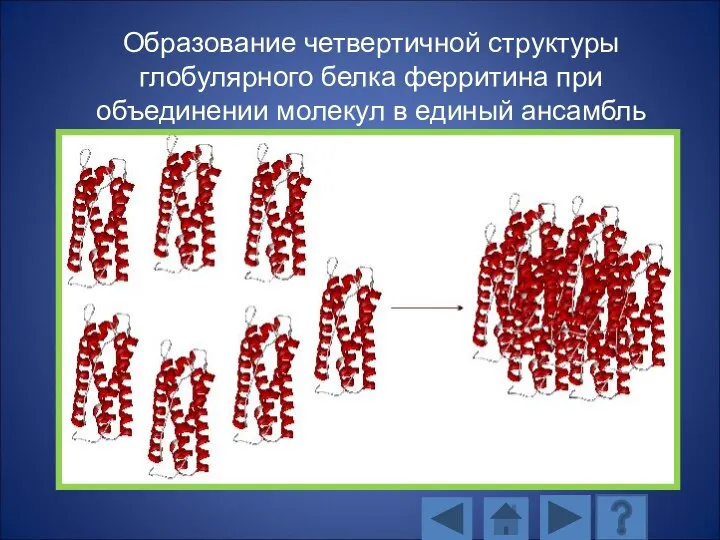
- 131. Образование четвертичной структуры глобулярного белка ферритина при объединении молекул в единый ансамбль
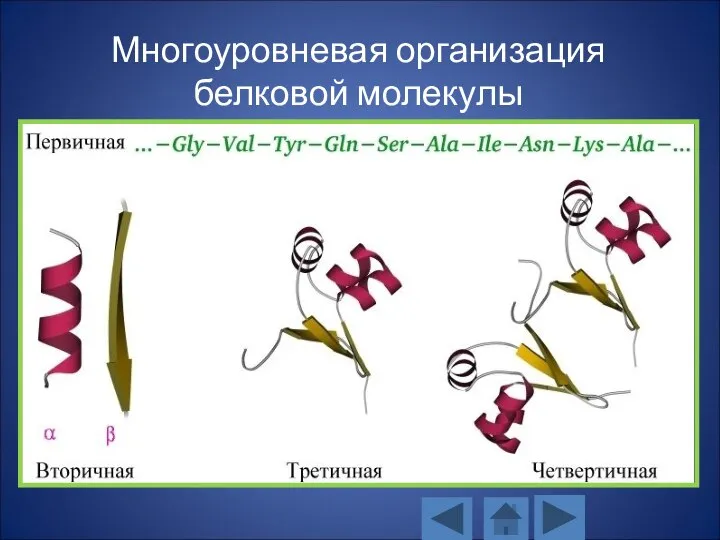
- 132. Многоуровневая организация белковой молекулы Первичная структура (Последовательность (цепочка) аминокислот) Вторичная структура (α-спираль, β-слой) Третичная структура (Трехмерная
- 133. Многоуровневая организация белковой молекулы
- 134. 2.3. Фолдинг Правильное сворачивание ( фолдинг ) полипептидных цепей некоторых белков в клетках эукариот обеспечивается специфическими
- 135. Шапероны обеспечивают: укладку новосинтезированные белков после выхода с рибосом в стабильную трехмерную структуру, укладку мультибелковых комплексов,
- 136. 3.1. Общие свойства Характерными физическими свойствами белков являются: высокая вязкость растворов, способность к набуханию, оптическая активность,
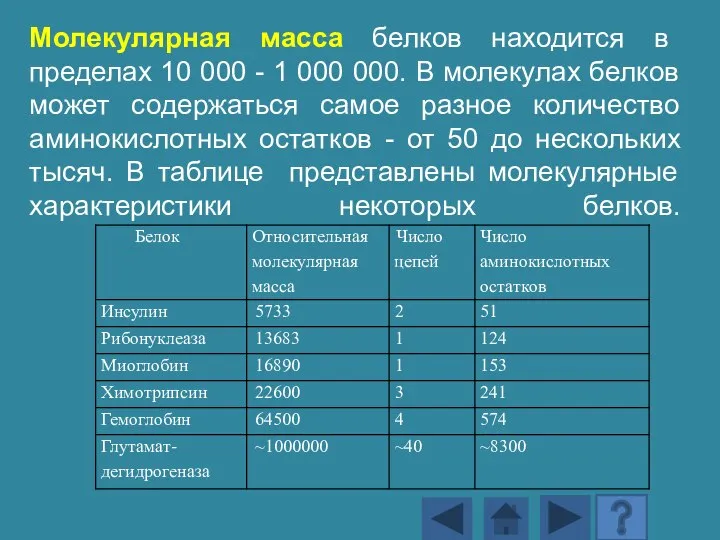
- 137. Молекулярная масса белков находится в пределах 10 000 - 1 000 000. В молекулах белков может
- 138. Сложность состава и высочайшую молекулярную массу белков можно подтвердить даже этой малой информацией:
- 139. НАТИВНОСТЬ - это уникальный комплекс физических, физико-химических, химических и биологических свойств белковой молекулы, который принадлежит ей,
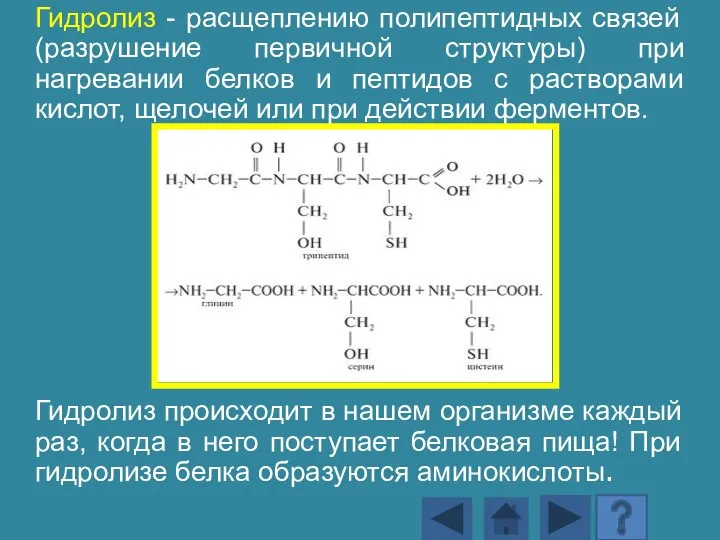
- 140. Гидролиз - расщеплению полипептидных связей (разрушение первичной структуры) при нагревании белков и пептидов с растворами кислот,
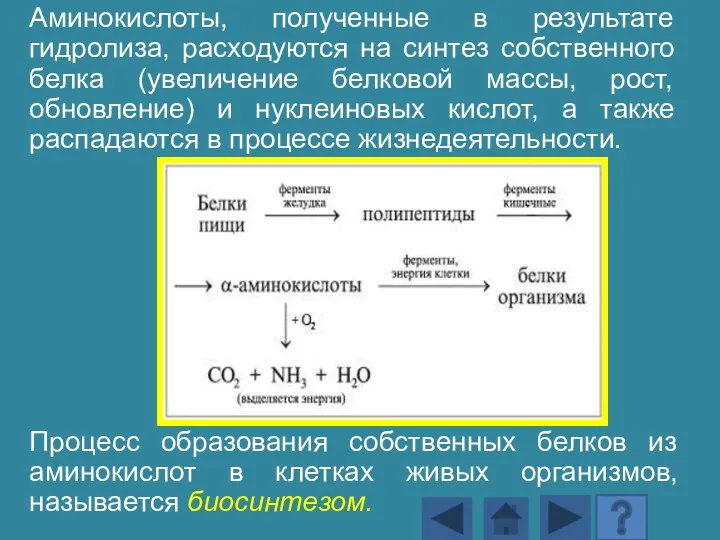
- 141. Аминокислоты, полученные в результате гидролиза, расходуются на синтез собственного белка (увеличение белковой массы, рост, обновление) и
- 142. 3.2.Электрические свойства белковых молекул Определяются присутствием на поверхности молекул белка положительно и отрицательно заряженных аминокислотных остатков.
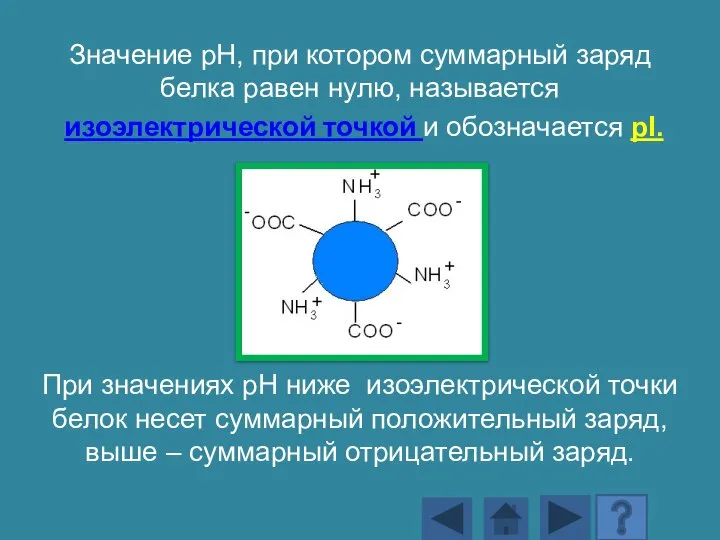
- 143. Значение рН, при котором суммарный заряд белка равен нулю, называется изоэлектрической точкой и обозначается pI. При
- 144. Изоэлектрическая точка для большинства белков находится в области рН от 4,5 до 6,5. Ниже приведены изоэлектрические
- 145. В изоэлектрической точке растворимость белка минимальна, так как его молекулы в таком состоянии электронейтральны и между
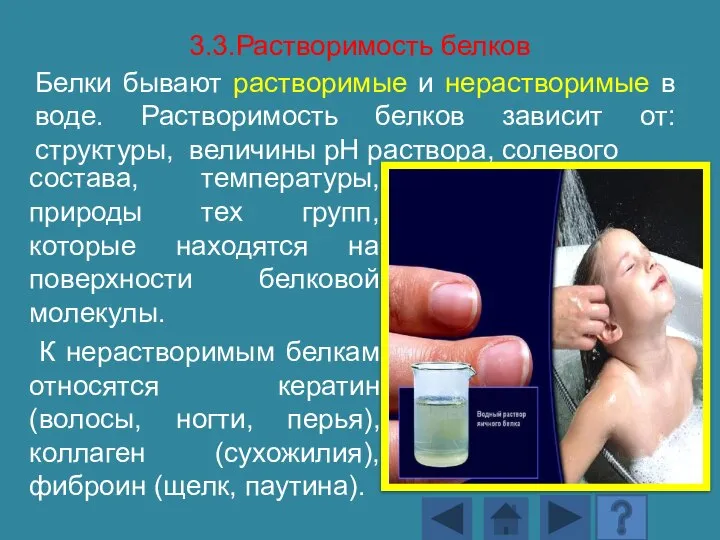
- 146. 3.3.Растворимость белков Белки бывают растворимые и нерастворимые в воде. Растворимость белков зависит от: структуры, величины рН
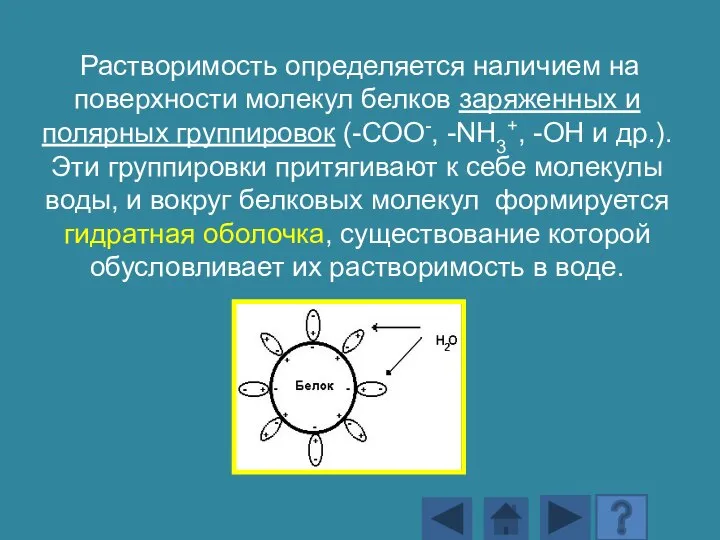
- 147. Растворимость определяется наличием на поверхности молекул белков заряженных и полярных группировок (-СОО-, -NH3+, -OH и др.).
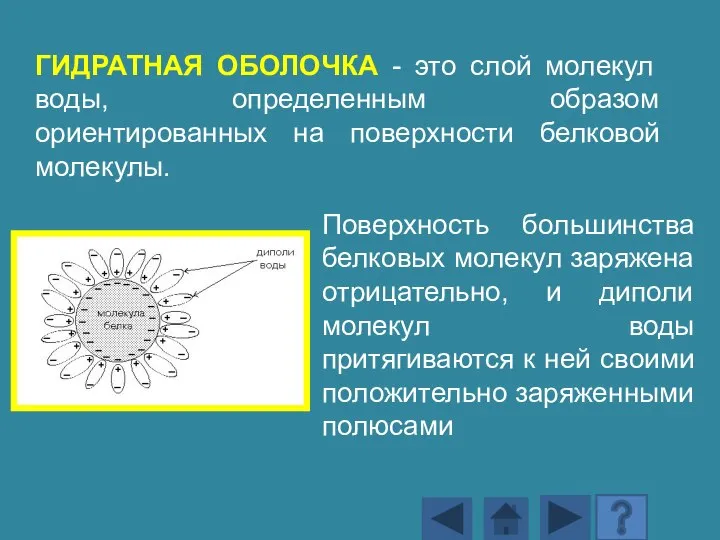
- 148. Поверхность большинства белковых молекул заряжена отрицательно, и диполи молекул воды притягиваются к ней своими положительно заряженными
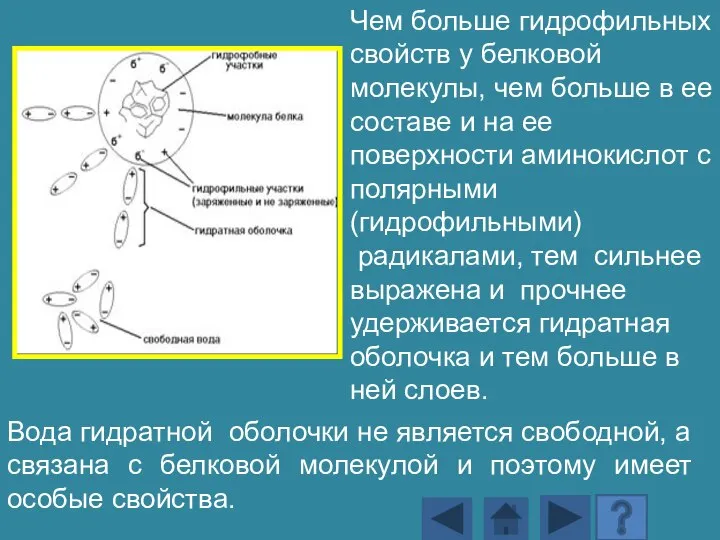
- 149. Чем больше гидрофильных свойств у белковой молекулы, чем больше в ее составе и на ее поверхности
- 150. Реакции осаждения делят на два вида. Обратимое осаждение белков предполагает выпадение белка в осадок под действием
- 151. Обратимое осаждение белков Для данного типа осаждения белков употребляют соли щелочных и щелочноземельных металлов (более часто
- 152. Осаждения с потерей нативных свойств белка - процесс необратимый. С белка снимается гидратная оболочка и заряд,
- 153. При кипячении белок выпадает в осадок. При нагревании до высоких температур усиливается тепловое движение молекул, ослабляются
- 154. Соли тяжелых металлов образуют соединения с SH-группами белков. Ядовиты для человека и животных. В медицинской практике
- 155. Существуют белки, которые растворяются только в растворах солей и не растворяются в чистой воде, такие белки
- 156. Водорастворимые белки образуют в воде коллоидные растворы. Эти растворы характеризуются высокой вязкостью, способностью рассеивать лучи видимого
- 157. Явление диализа лежит в основе действия аппарата "искусственная почка", который широко используется в медицине для лечения
- 158. При определенных условиях белки могут образовывать гели. При образовании геля молекулы белка формируют густую сеть, внутреннее
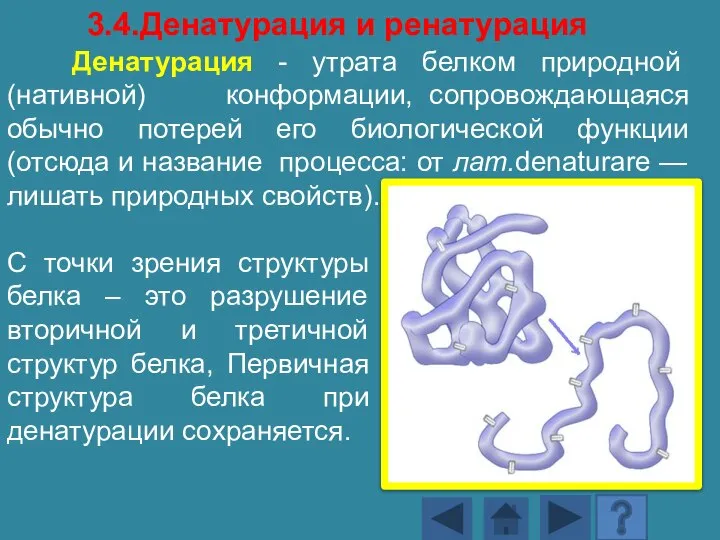
- 159. Денатурация - утрата белком природной (нативной) конформации, сопровождающаяся обычно потерей его биологической функции (отсюда и название
- 160. Факторы, вызывающие денатурацию Физические факторы Высокие температуры. Для разных белков характерна различная чувствительность к тепловому воздействию.
- 161. Денатурация может быть обратимой (ренатурация) и необратимой. Самый известный случай денатурации белка в быту — это
- 162. Денатурация белков происходит в желудке, где имеется сильнокислая среда (рН 0,5 - 1,5), и это способствует
- 163. Ренатурация (обратимая денатурация) – восстановление физико-химических и биологических свойств белка (нативной структуры) при снятии денатурирующего фактора.
- 164. При денатурации белков в клетках, в живом организме вырабатываются специфические белки шапероны, которые: - «узнают» частично
- 165. 3.5. Качественные (цветные) реакции на белки
- 166. 3.5. Качественные реакции на белки Биуретовая реакция – фиолетовое окрашивание при действии солей меди (II) в
- 167. Реакция обязана наличию пептидных связей в белках а также продуктах их гидролиза – пептидах: Интенсивность окраски
- 168. Нингидриновая реакция Сущность реакции состоит в образовании соединения, окрашенного в сине-фиолетовый цвет, состоящего из нингидрина и
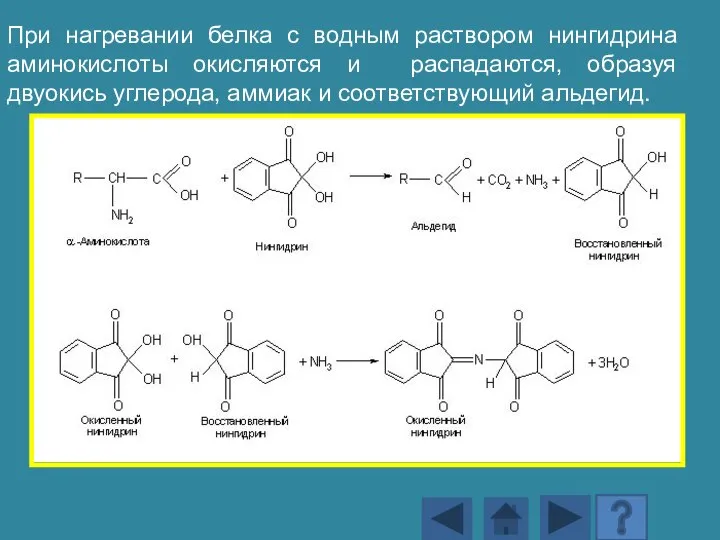
- 169. При нагревании белка с водным раствором нингидрина аминокислоты окисляются и распадаются, образуя двуокись углерода, аммиак и
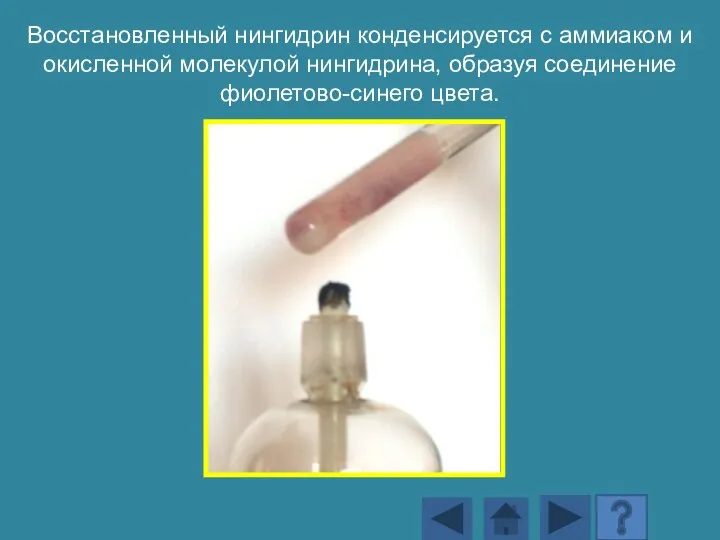
- 170. Восстановленный нингидрин конденсируется с аммиаком и окисленной молекулой нингидрина, образуя соединение фиолетово-синего цвета.
- 171. Интенсивность окраски полученного сине-фиолетового комплекса (пурпур Руэманна), пропорциональна количеству α-аминокислот. Желатин Яичный альбумин
- 172. Ксантопротеиновая реакция – появление желтого окрашивания при действии концентрированной азотной кислоты на белки, содержащие остатки ароматических
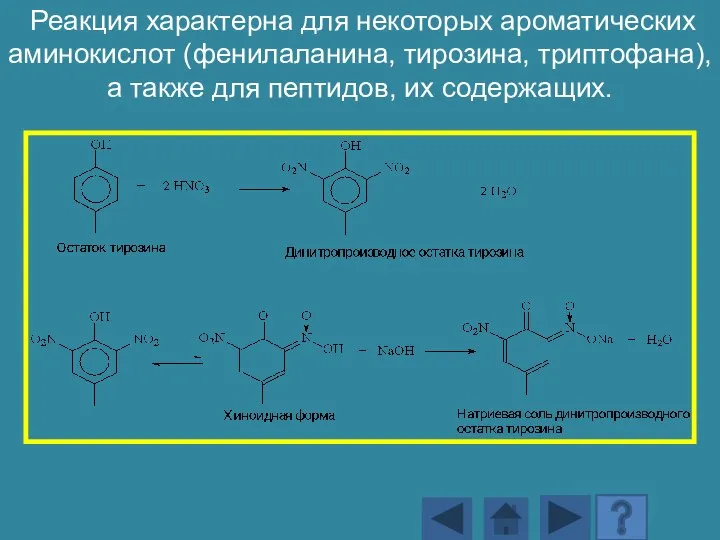
- 173. Реакция характерна для некоторых ароматических аминокислот (фенилаланина, тирозина, триптофана), а также для пептидов, их содержащих.
- 174. При действии азотной кислоты образуется нитросоединение желтого цвета. Нитропроизводные могут реагировать со щелочью с образованием натриевой
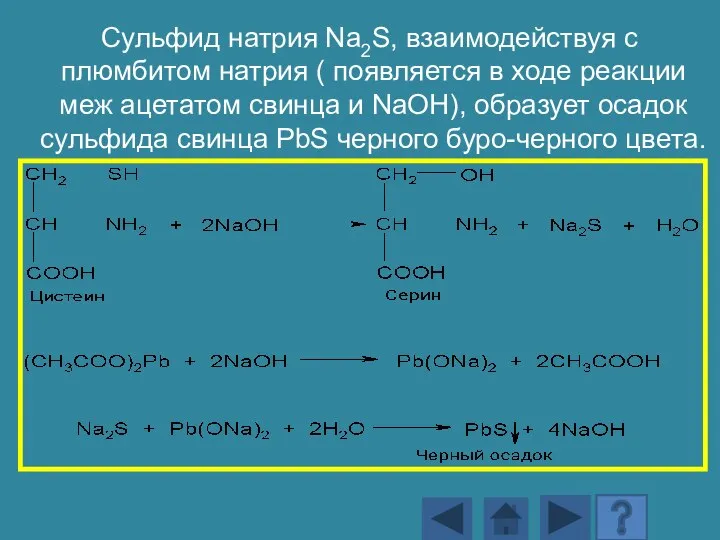
- 175. Реакция Фоля Реакция на серусодержащие аминокислоты, в ходе которой появляется черное либо бурое окрашивание сульфида свинца
- 176. Сульфид натрия Na2S, взаимодействуя с плюмбитом натрия ( появляется в ходе реакции меж ацетатом свинца и
- 177. Тепловая денатурация белка При нагревании белки денатурируют. На процесс денатурации оказывают сильное влияние рН раствора и
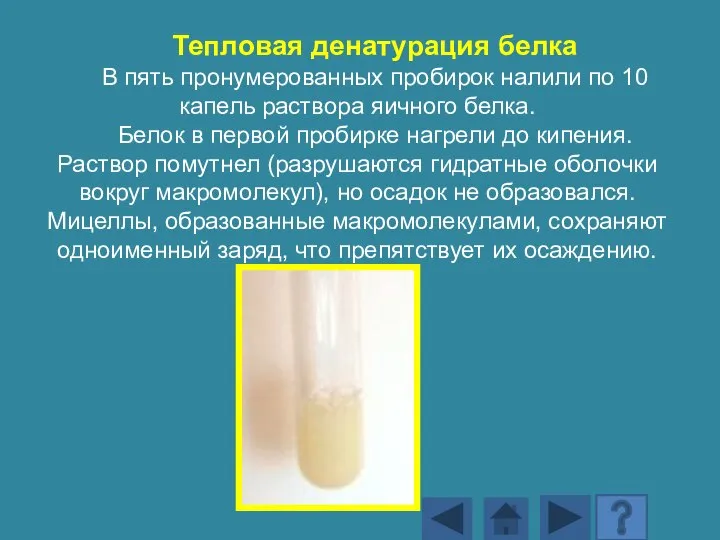
- 178. Тепловая денатурация белка В пять пронумерованных пробирок налили по 10 капель раствора яичного белка. Белок в
- 179. Тепловая денатурация белка К раствору белка во второй пробирке добавили одну каплю 1 %-го раствора уксусной
- 180. Тепловая денатурация белка К раствору белка в третьей пробирке прибавили 1 – 2 капли 10 %-го
- 181. Тепловая денатурация белка В четвертую пробирку добавили 1 – 2 капли 10 %-го раствора гидроксида натрия
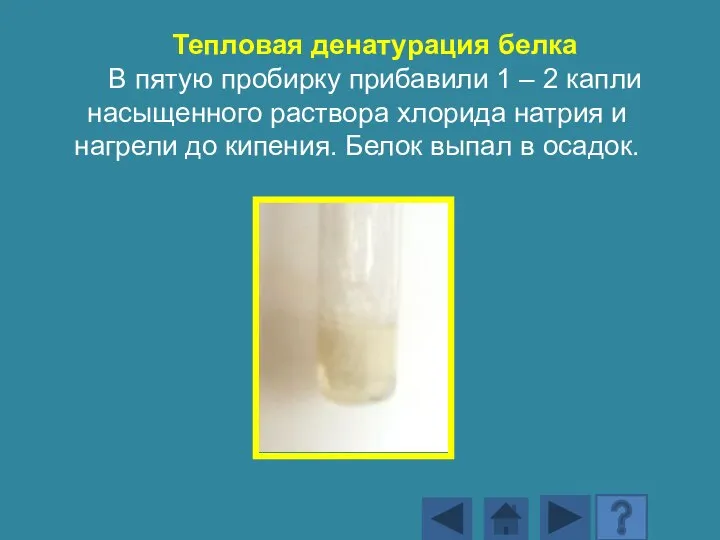
- 182. Тепловая денатурация белка В пятую пробирку прибавили 1 – 2 капли насыщенного раствора хлорида натрия и
- 183. Интересно знать! Прочность белковых молекул просто поразительна! Человеческий волос прочнее меди и может соперничать со специальными
- 184. Белки могут выполнять все известные в природе функции, кроме одной - синтезировать сами себя. Основные функции
- 185. 4.1. Ферментативная функция Самый многообразный и наиболее специализированный класс белков составляют ферменты – белки, обладающие каталитической
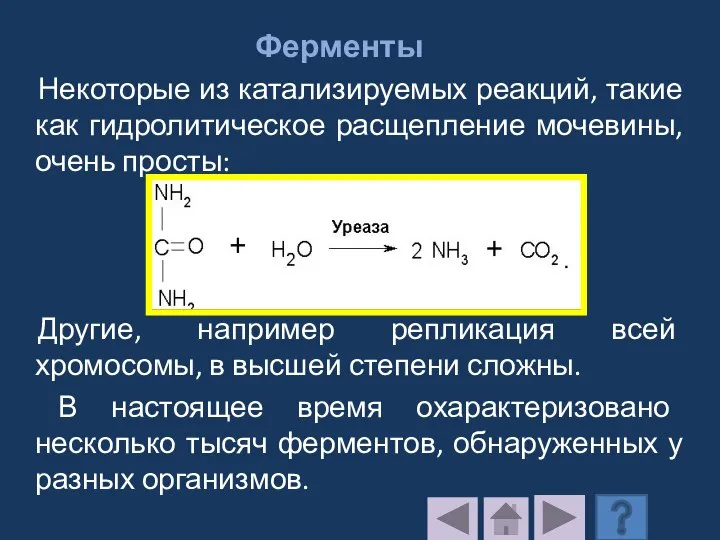
- 186. Ферменты Некоторые из катализируемых реакций, такие как гидролитическое расщепление мочевины, очень просты: Другие, например репликация всей
- 187. Катализирует преквращение этанола и других алифатических спиртов. Алкогольдегидрогеназа
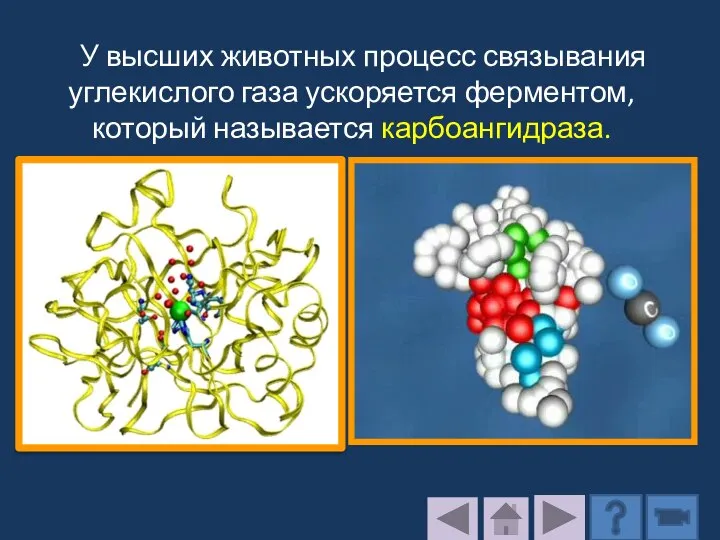
- 188. У высших животных процесс связывания углекислого газа ускоряется ферментом, который называется карбоангидраза.
- 189. В процессе пищеварения участвуют группы ферментов катализирующие расщепление определенных веществ: – амилазы, катализирующие расщепление крахмала на
- 190. Интересно знать! Созданы препараты содержащие ферменты поджелудочной железы. Например, мезим-форте®, который содержит 4200 ЕД амилазы, 3500
- 191. Коллаген Фибриллярный белок. Содержится в соединительной ткани: в сухожилиях, костях, хрящах, стенках сосудов. Способен набухать в
- 192. Интересно знать! Коллаген является наиболее известным препаратом для коррекции морщин и других дефектов кожи. Содержание собственного
- 193. Кератин Нерастворимый прочный белок. Содержится в волосах, рогах, копытах, шерсти, ногтях, панцире, перьях.
- 194. Структурные белки Эластин Cтруктурный белок способный растягиваться в двух измерениях. Содержится в связках, стенках капилляров и
- 195. Фиброин Главный компонент шелковых нитей и паутины. Имеет мягкие подвижные волокна. Обладает структурой антипараллельного складчатого листа
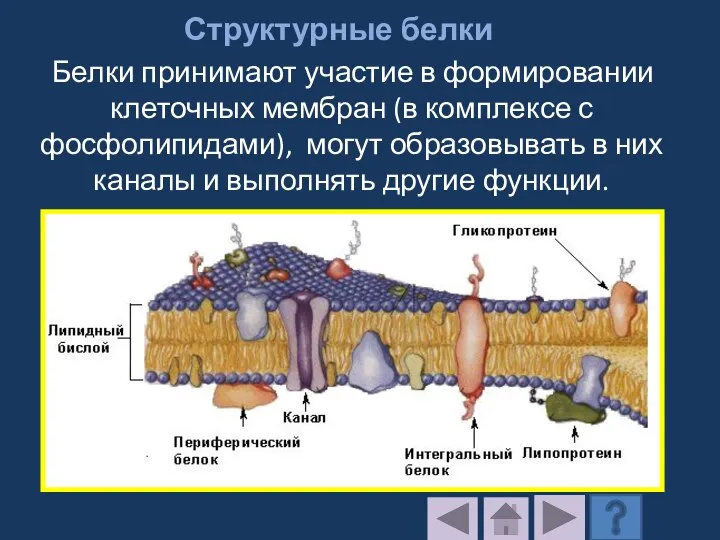
- 196. Структурные белки Белки принимают участие в формировании клеточных мембран (в комплексе с фосфолипидами), могут образовывать в
- 197. 4.3. Транспортная функция Транспортные белки переносят специфические молекулы и ионы. Внутрь клетки через биологическую мембрану должны
- 198. Бактериородопсин проводит протон через мембрану. Сидящие на a-спиралях бактериородопсина гидрофобные группы обращены "наружу", к (тоже гидрофобным)
- 199. У многоклеточных организмов существует система транспорта веществ от одних органов к другим. Например, в крови позвоночных
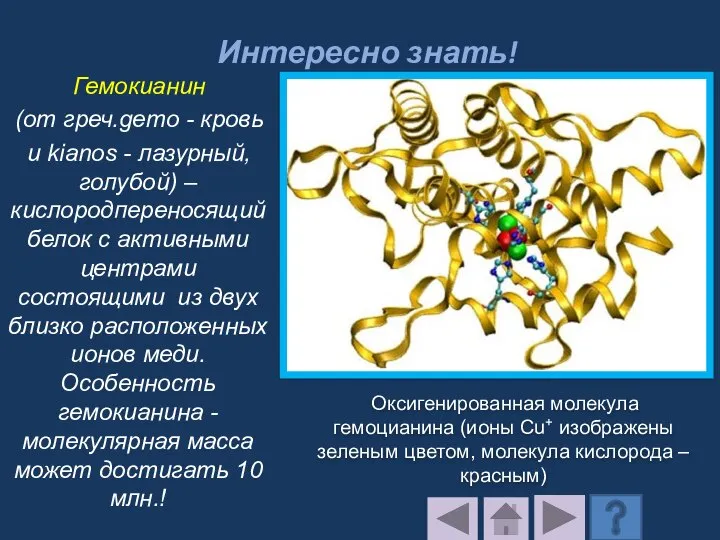
- 200. Интересно знать! Гемокианин (от греч.gemo - кровь и kianos - лазурный,голубой) –кислородпереносящий белок с активными центрами
- 201. Интересно знать! Кислород переносится «голубой кровью» у кальмаров, омаров, крабов, осьминогов, каракатиц, улиток, пауков.
- 202. Транспортные белки В плазме крови многоклеточных организмов находится транспортный белок - сывороточный альбумин, который обладает способностью
- 203. В семенах многих растений запасены пищевые белки, потребляемые на первых стадиях развития зародыша. К запасным белкам
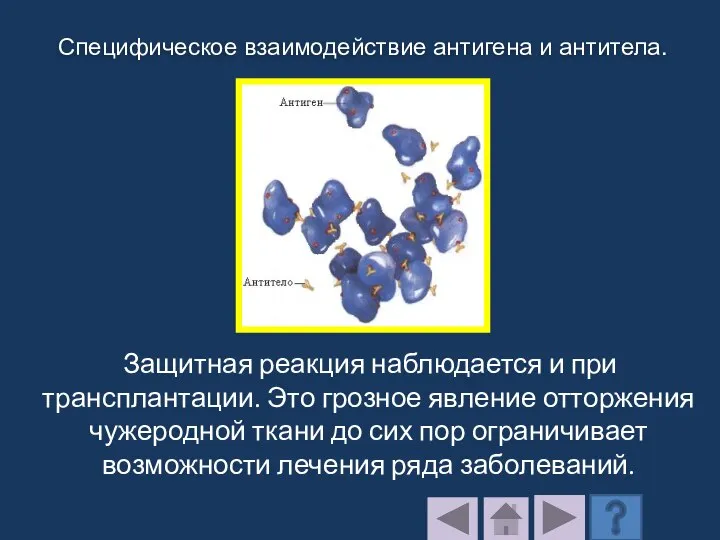
- 204. К этой группе относятся белки, защищающие организм от вторжения других организмов или предохраняющие его от повреждений.
- 205. Защитная реакция наблюдается и при трансплантации. Это грозное явление отторжения чужеродной ткани до сих пор ограничивает
- 206. Фибрин - нерастворимый белок, образующийся в процессе свертывания крови. Нити фибрина, полимеризуясь, образуют основу тромба, останавливающего
- 207. Стрессовые белки синтезируются в растениях в ответ на различные экстремальные воздействия: окислительный стресс, повышение и понижение
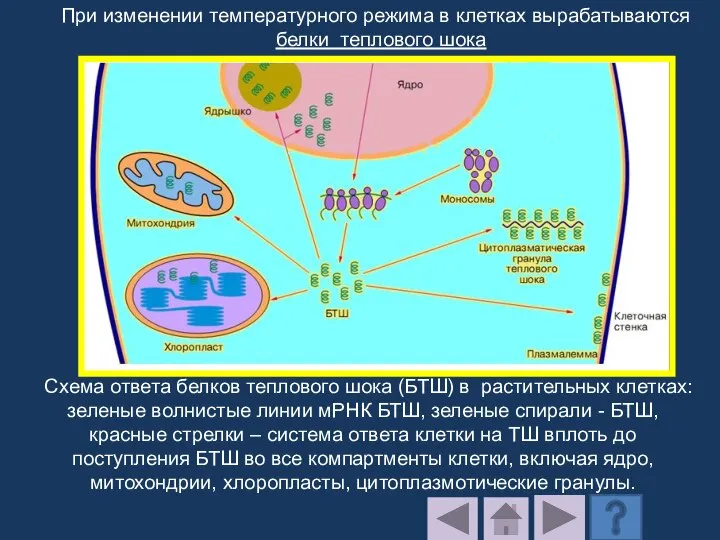
- 208. Схема ответа белков теплового шока (БТШ) в растительных клетках: зеленые волнистые линии мРНК БТШ, зеленые спирали
- 209. Белки - рецепторы «Благоденствие» и выживание каждой клетки и организма в целом зависят от того, насколько
- 210. Белки - рецепторы Существует три основных типа рецепторов, интегрированных во внешнюю клеточную мембрану: рецепторы, сопряженные с
- 211. Рецепторы, сопряженные с G–белками G- белки – это регуляторы, включающие или выключающие активность других молекул. Когда
- 212. Структура холинэргического рецептора никотинового типа, формирующего ионный канал. Субъединицы полипептидной цепи пронизывают 4 раза липидный бислой.
- 213. Рецепторы, ассоциированные с ферментативной активностью Белки-рецепторы данного типа могут обладать собственной активностью, которая проявляется при связывании
- 214. Некоторые белки наделяют клетку или организм способностью сокращаться, изменять форму или передвигаться. Сокращение мышцы обеспечивается скольжение
- 215. Белки обладают чрезвычайно широкой биологической активностью. Они регулируют работу сердечно-сосудистой, иммунной, половой, эндокринной, пищеварительной и других
- 216. К ним принадлежат некоторые гормоны, вырабатываемые в железах внутренней секреции. Инсулин и глюкагон регулируют уровень глюкозы
- 217. Окситоцин человека Пептидные гормоны Окситоцин Стимулирует сокращение матки у беременных самок. Стимулирует выделение молока из молочных
- 218. Вазопрессин человека Пептидные гормоны Вазопрессин Обладает сильным антидиуретическим действием (регулирует водный обмен), стимулируя реабсорбцию воды почками.
- 219. Гормон роста (соматотропин) определяет размеры тела, секретируется передней долей гипофиза. Гормоном роста соматотропин называют за то,
- 220. Недостаток гормона роста в детском возрасте связан в основном с генетическими дефектами и вызывает задержку роста
- 221. 4.9. Токсины Большая группа фармакологически активных пептидов, в том числе брадикинин, физалемин, церулеин и др., найдены
- 222. Некоторые из них вызывают быстрое сокращение внесосудистой мускулатуры и получили название тахикининов. Брадикинин Арг-Про-Про-Гли-Фен-Сер-Про-Фен-Арг Ранакинин R
- 223. Токсины Мелиттин — основной компонент пчелиного яда, содержание его в целом яде достигает 50%. Это сильный
- 224. токсины Все токсические полипептиды змей делят на: - токсины, специфически блокирующие Н-холинорецепторы субсинаптической мембраны скелетной мускулатуры
- 225. токсины Бледная поганка Смертельно ядовитый гриб. α-аманитин
- 226. Интересно знать! Нобелевская премия по химии 2008 года присуждена трём биологам — японцу Осаму Шимомуре и
- 227. Белки используют в промышленности для окрашивания тканей, при рафинировании сахара, в различных операциях по фотографии
- 228. Интересно знать! В плазме некоторых антарктических рыб содержатся белки со свойствами антифриза, предохраняющие рыб от замерзания,
- 229. Кроме выше рассмотренных функций, белки участвуют в экспрессии генетической информации, передаче нервных импульсов, поддерживают онкотическое давление
- 230. Белки как необходимые компоненты пищи Необходимые для человека белки содержат продукты питания различного происхождения. Животные продукты
- 231. Содержание белка в животных продуктах Мясо, рыба и птица усваиваются организмом в разной степени и поэтому
- 232. Содержание белка в растительных продуктах Бобовые и крупы отличаются особенно высоким содержанием белка. Чечевица, соя и
- 233. Дефицит белка в питании уменьшает устойчивость организма к инфекциям, так как снижается уровень образования антител. Нарушается
- 234. Заключение Сегодня на «коротком» пути от гена к белку (он весь умещается в пределах одной клетки)
- 235. После того, как синтезирована аминокислотная последовательность белка, получившийся продукт еще не стал белком в химическом и
- 236. Пространственную структуру белка сегодня можно определить лишь экспериментально - с помощью рентгеноструктурного анализа кристаллов белка или
- 238. Скачать презентацию
Слайд 2СОДЕРЖАНИЕ
Ведение
1. Общая характеристика белковых молекул
1.1. Элементный состав белков
1.2. Аминокислотный состав белков
1.2.1. Характеристика
СОДЕРЖАНИЕ
Ведение
1. Общая характеристика белковых молекул
1.1. Элементный состав белков
1.2. Аминокислотный состав белков
1.2.1. Характеристика

Слайд 33.Физико-химические свойства белков
3.1. Общие свойства
3.2. Электрические свойства белковых молекул
3.3. Растворимость белков
3.4. Денатурация
3.Физико-химические свойства белков
3.1. Общие свойства
3.2. Электрические свойства белковых молекул
3.3. Растворимость белков
3.4. Денатурация

Слайд 4Условные обозначения
Глицин
Гиперссылка
Глоссарий
Шаг назад
Шаг вперед
К содержанию
Видеофрагмент
-
-
-
-
-
-
Условные обозначения
Глицин
Гиперссылка
Глоссарий
Шаг назад
Шаг вперед
К содержанию
Видеофрагмент
-
-
-
-
-
-
Слайд 5
Ведение
Белки или протеины
(от греч. protos–первый, важнейший) –
это высокомолекулярные азотсодержащие органические
Ведение
Белки или протеины
(от греч. protos–первый, важнейший) –
это высокомолекулярные азотсодержащие органические

Слайд 6Кроме понятия «белок», в химии встречается термины «пептид» и «полипептид». Пептидом обычно
Кроме понятия «белок», в химии встречается термины «пептид» и «полипептид». Пептидом обычно

Слайд 7 Совокупность белковых молекул имеющихся в организме называется протеомом. Термин "протеом" предложил
Совокупность белковых молекул имеющихся в организме называется протеомом. Термин "протеом" предложил

Слайд 8 Основные химические элементы, входящие в состав белков
С(углерод) – 50-55%
O(кислород) – 21-23%
H(водород) –
Основные химические элементы, входящие в состав белков
С(углерод) – 50-55%
O(кислород) – 21-23%
H(водород) –

Слайд 91.2. Аминокислотный состав белков
Белки построены в основном двадцатью стандартными (основными, природными, белковыми,
1.2. Аминокислотный состав белков
Белки построены в основном двадцатью стандартными (основными, природными, белковыми,

Слайд 10 Аминокислоты входящие в состав белка являются
α-аминокислотами и представляют собой
производные карбоновых
Аминокислоты входящие в состав белка являются
α-аминокислотами и представляют собой
производные карбоновых

Слайд 11Общая формула α-аминокислот имеет следующий вид:
R - боковые цепочки (радикалы), не
Общая формула α-аминокислот имеет следующий вид:
R - боковые цепочки (радикалы), не
Слайд 12Природные аминокислоты имеют тривиальные названия, которые происходят иногда от источника из которого
Природные аминокислоты имеют тривиальные названия, которые происходят иногда от источника из которого

Слайд 131.2.1. Характеристика стандартных аминокислот
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми
1.2.1. Характеристика стандартных аминокислот
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми

Слайд 14Структурная формула
Пространственное строение радикала
(Серыми кружками показаны атомы С, светло-серыми — H атомы
Структурная формула
Пространственное строение радикала
(Серыми кружками показаны атомы С, светло-серыми — H атомы

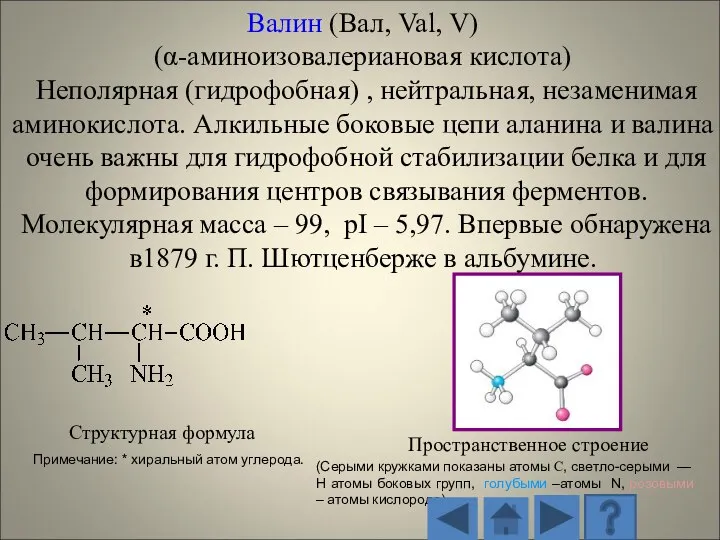
Слайд 15Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Слайд 16Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

Слайд 17Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

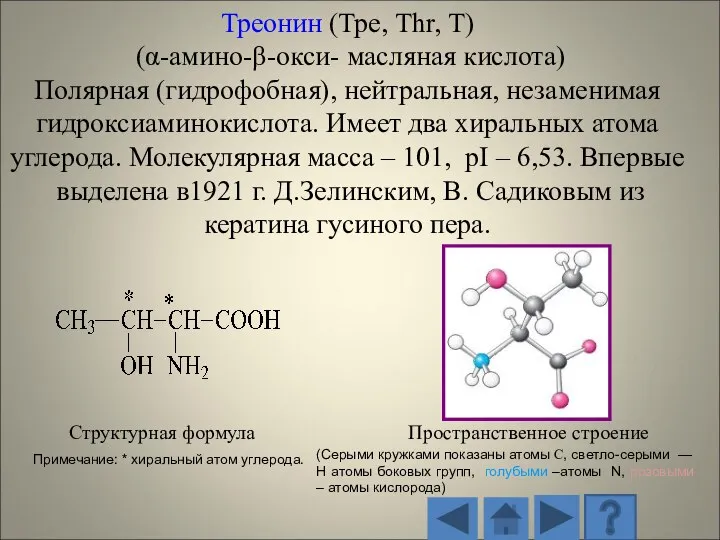
Слайд 18Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Слайд 19Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

Слайд 20Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
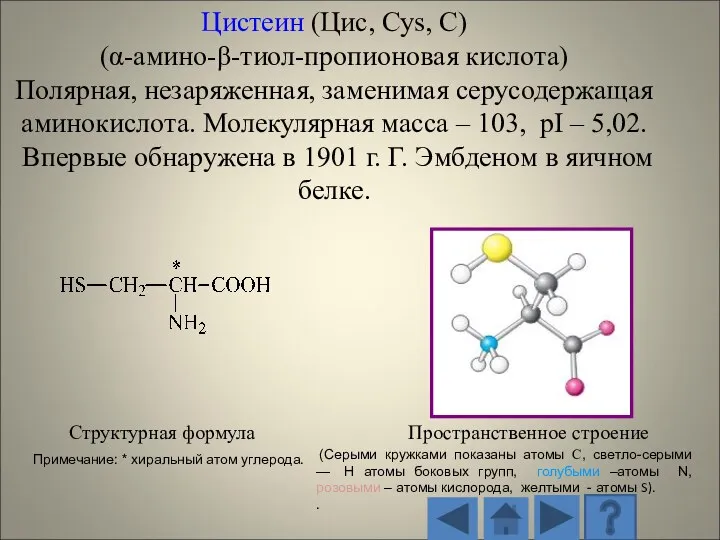
Слайд 21Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
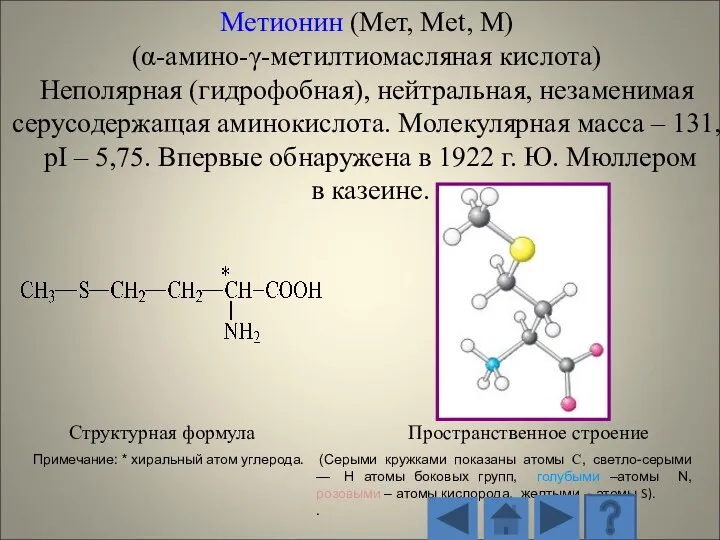
Слайд 22Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
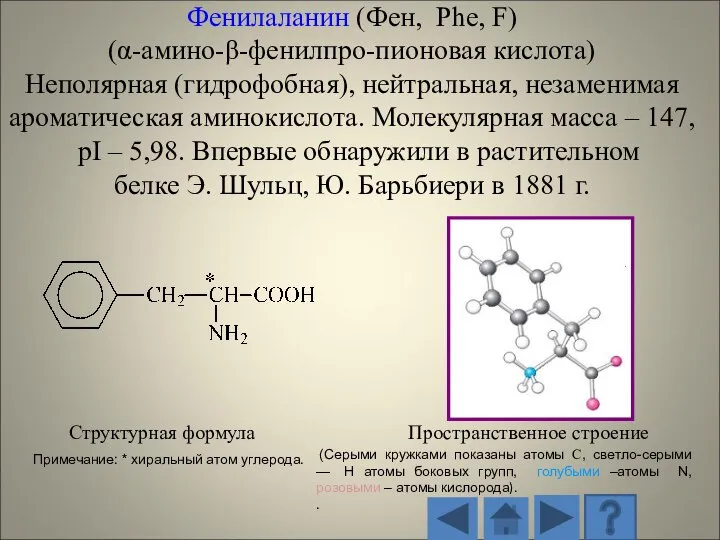
Слайд 23Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
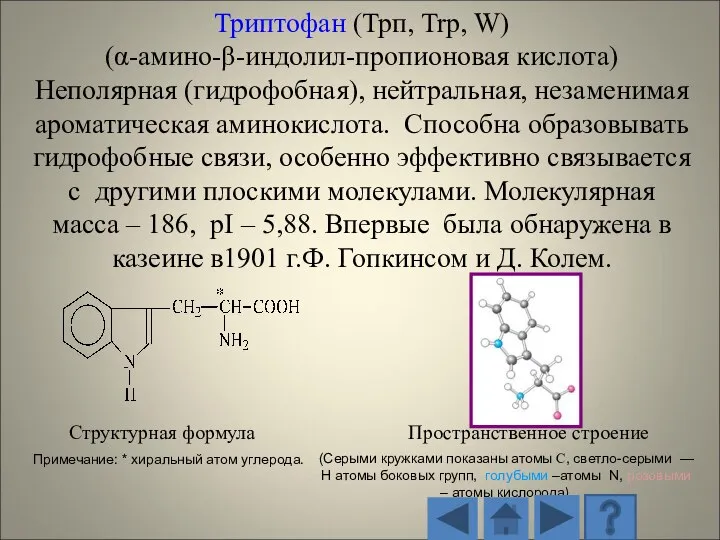
Слайд 24Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
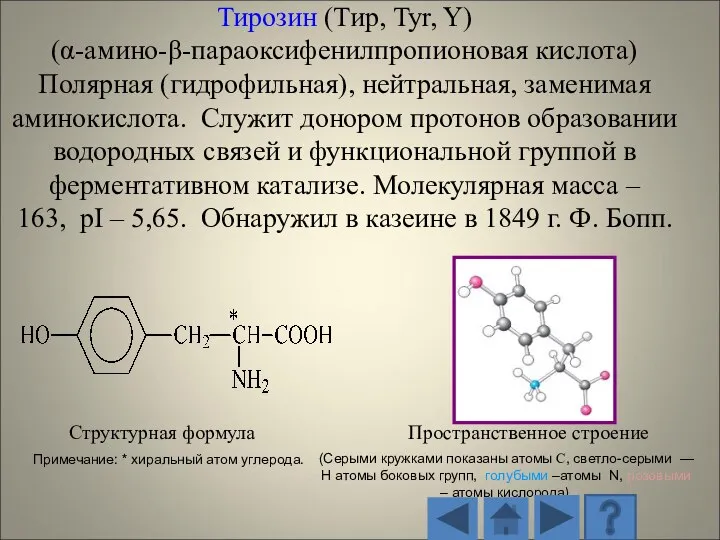
Слайд 25Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
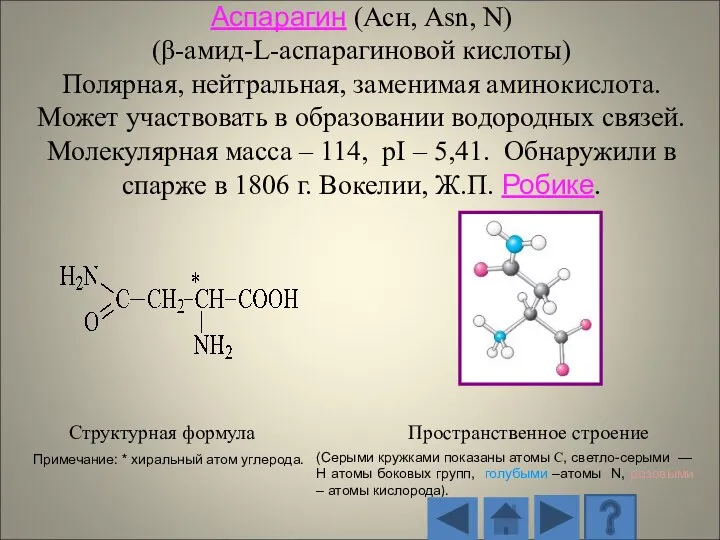
Слайд 26Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

Слайд 27Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

Слайд 28Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы

Слайд 29Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

Слайд 30Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

Слайд 31Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых

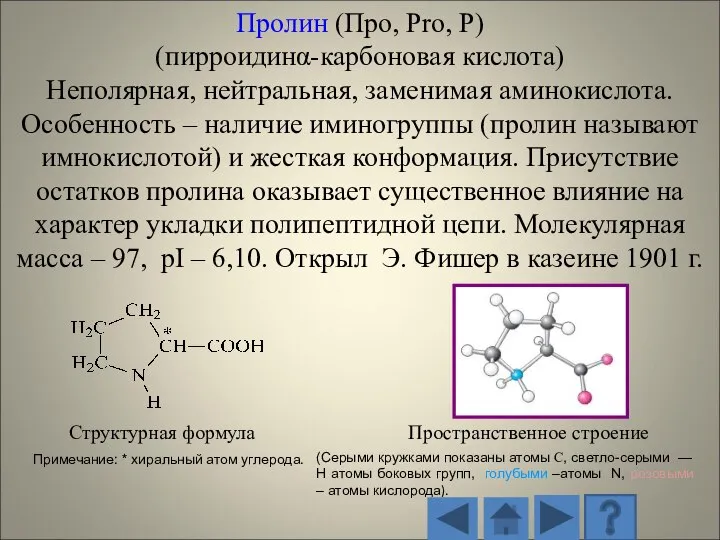
Слайд 32Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
Структурная формула
Пространственное строение
(Серыми кружками показаны атомы С, светло-серыми — H атомы боковых
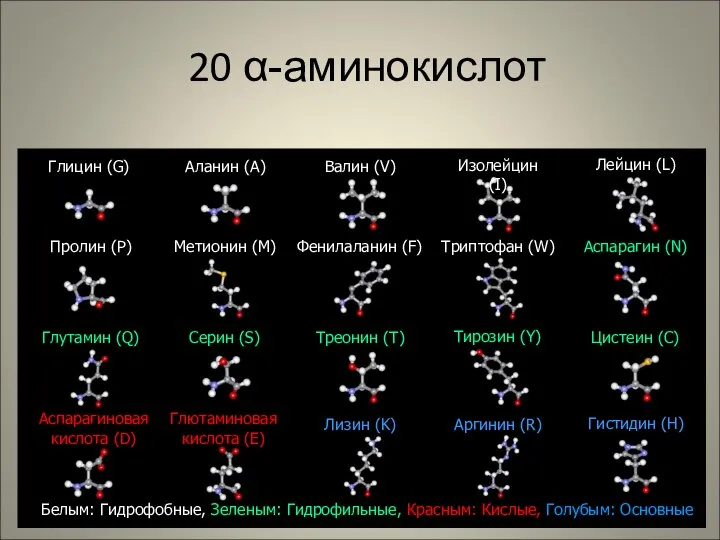
Слайд 3320 α-аминокислот
Глицин (G)
Глютаминовая кислота (E)
Аспарагиновая кислота (D)
Метионин (M)
Треонин (T)
Серин (S)
Глутамин (Q)
Аспарагин (N)
Триптофан
20 α-аминокислот
Глицин (G)
Глютаминовая кислота (E)
Аспарагиновая кислота (D)
Метионин (M)
Треонин (T)
Серин (S)
Глутамин (Q)
Аспарагин (N)
Триптофан
Слайд 34 Важным свойством аминокислот является оптическая изомерия. Это свойство связано с наличием в
Важным свойством аминокислот является оптическая изомерия. Это свойство связано с наличием в

Слайд 35 Для обозначения изомеров используется
D, L-номенклатура.
Например, для аланина проекции Фишера обоих изомеров
Для обозначения изомеров используется
D, L-номенклатура.
Например, для аланина проекции Фишера обоих изомеров
Слайд 36 В числе белковых аминокислот имеются две аминокислоты (треонин и изолейцин), которые содержат
В числе белковых аминокислот имеются две аминокислоты (треонин и изолейцин), которые содержат
Слайд 37L - и D-изомеры могут взаимно превращаться друг в друга. Этот процесс
L - и D-изомеры могут взаимно превращаться друг в друга. Этот процесс

Слайд 38В белке зубов – дентине – L-аспарагиновая кислота самопроизвольно рацемизуется при температуре
В белке зубов – дентине – L-аспарагиновая кислота самопроизвольно рацемизуется при температуре

Слайд 39
1.2.3. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА АМИНОКИСЛОТ
Физические свойства
Аминокислоты представляют собой кристаллические вещества с высокими (выше
1.2.3. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА АМИНОКИСЛОТ
Физические свойства
Аминокислоты представляют собой кристаллические вещества с высокими (выше

Слайд 40Химические свойства
Аминокислоты содержат амино- и карбоксильную группы. В силу этого они
Химические свойства
Аминокислоты содержат амино- и карбоксильную группы. В силу этого они

Слайд 41Химические свойства
При растворении аминокислоты в воде, например, глицина, его карбоксильная группа диссоциирует
Химические свойства
При растворении аминокислоты в воде, например, глицина, его карбоксильная группа диссоциирует

Слайд 42
Кислотно-основные свойства
не ионная форма;
идеализированная аминокислота
цвиттер-ион
Кислотно-основные свойства
не ионная форма;
идеализированная аминокислота
цвиттер-ион

Слайд 43Кислотно-основные свойства
В кислой среде аминокислота, присоединяя ион водорода, образует катион:
В
Кислотно-основные свойства
В кислой среде аминокислота, присоединяя ион водорода, образует катион:
В

Слайд 44Важной особенностью аминокислот является их способность к поликонденсации, приводящей к образованию полиамидов,
Важной особенностью аминокислот является их способность к поликонденсации, приводящей к образованию полиамидов,

Слайд 45Для α-аминокислот характерны следующие химические реакции:
реакции по карбоксильной группе;
реакции по
Для α-аминокислот характерны следующие химические реакции:
реакции по карбоксильной группе;
реакции по

Слайд 46
1.2.4.Классификация α-аминокислот
В зависимости от подхода можно выделить следующие типы классификаций аминокислот:
1.2.4.Классификация α-аминокислот
В зависимости от подхода можно выделить следующие типы классификаций аминокислот:

Слайд 47Химическая классификация
В соответствии с химической структурой радикала аминокислоты делят на алифатические,
В соответствии с химической структурой радикала аминокислоты делят на алифатические,

Слайд 48Алифатические
Моноаминомонокарбоновые: аланин, валин, глицин, изолейцин, лейцин.
Оксимоноаминокарбоновые: серин, треонин.
Моноаминодикарбоновые: аспарагиновая
Алифатические
Моноаминомонокарбоновые: аланин, валин, глицин, изолейцин, лейцин.
Оксимоноаминокарбоновые: серин, треонин.
Моноаминодикарбоновые: аспарагиновая

Слайд 49Ароматические: фенилаланин, тирозин
- Гетероциклические: триптофан, гистидин, пролин (также входит в группу
Ароматические: фенилаланин, тирозин
- Гетероциклические: триптофан, гистидин, пролин (также входит в группу

Слайд 50Физико-химическая классификация
основана на полярности радикалов
(R-групп), т.е. способности их к взаимодействию
основана на полярности радикалов
(R-групп), т.е. способности их к взаимодействию

Слайд 51 Выделяют четыре класса аминокислот
(в зависимости от типа радикала) :
Выделяют четыре класса аминокислот
(в зависимости от типа радикала) :
Слайд 52К неполярным (гидрофобным) относятся аминокислоты с неполярными алифатическими (аланин, валин, лейцин, изолейцин)
К неполярным (гидрофобным) относятся аминокислоты с неполярными алифатическими (аланин, валин, лейцин, изолейцин)

Слайд 53Полярные незаряженные аминокислоты в сравнении с неполярными лучше растворяются в воде, более
Полярные незаряженные аминокислоты в сравнении с неполярными лучше растворяются в воде, более

Слайд 54К положительно заряженным аминокислотам принадлежат лизин, гистидин и аргинин, в ионизированном виде
К положительно заряженным аминокислотам принадлежат лизин, гистидин и аргинин, в ионизированном виде

Слайд 55 К отрицательно заряженным аминокислотам относятся аспарагиновая и глутаминовая кислоты. Они содержат
К отрицательно заряженным аминокислотам относятся аспарагиновая и глутаминовая кислоты. Они содержат

Слайд 56 Биологическая классификация
основана на способности
аминокислот, принимающие участие в построении белков
Биологическая классификация
основана на способности
аминокислот, принимающие участие в построении белков

Слайд 57 Заменимые аминокислоты могут синтезироваться в организме человека
(их 10).
Незаменимые
Заменимые аминокислоты могут синтезироваться в организме человека
(их 10).
Незаменимые


Слайд 59
Незаменимые аминокислоты входят часто в состав пищевых добавок, используются в качестве
Незаменимые аминокислоты входят часто в состав пищевых добавок, используются в качестве

Слайд 60C 1981 г. в США разрешено использование низкокалорийной добавки для придания продуктам
C 1981 г. в США разрешено использование низкокалорийной добавки для придания продуктам

Слайд 61Аргинин содержится в твороге, морепродуктах,грецких орехах,шоколаде.
Валин содержится в зерновых, мясе, грибах, арахисе,
Аргинин содержится в твороге, морепродуктах,грецких орехах,шоколаде.
Валин содержится в зерновых, мясе, грибах, арахисе,

Слайд 62Интересно знать!
Исключительно важную роль играет сбалансированность питания человека по аминокислотам. При
Интересно знать!
Исключительно важную роль играет сбалансированность питания человека по аминокислотам. При

Слайд 631.2.5. Производные аминокислот
Нестанда́ртные аминокисло́ты — аминокислоты, которые, хотя и не входят в
1.2.5. Производные аминокислот
Нестанда́ртные аминокисло́ты — аминокислоты, которые, хотя и не входят в

Слайд 64 Среди таких превращений следует в первую очередь отметить образование дисульфидных мостиков
Среди таких превращений следует в первую очередь отметить образование дисульфидных мостиков
Слайд 65 Важным примером модификации аминокислотных остатков является превращение остатков пролина в остатки
Важным примером модификации аминокислотных остатков является превращение остатков пролина в остатки
Слайд 66Еще одним весьма важным видом модификации белков является фосфорилирование гидроксогрупп остатков серина,
Еще одним весьма важным видом модификации белков является фосфорилирование гидроксогрупп остатков серина,

Слайд 67
По составу белковой молекулы
1.3. Простые и сложные белки
По составу белковой молекулы
1.3. Простые и сложные белки
Слайд 68Простые белки
Простые белки состоят только из аминокислотных остатков и не содержат других
Простые белки
Простые белки состоят только из аминокислотных остатков и не содержат других

Слайд 69Альбумины (от лат. albumen, род. падеж albuminis - белок) - водорастворимые глобулярные
Альбумины (от лат. albumen, род. падеж albuminis - белок) - водорастворимые глобулярные

Слайд 70Сывороточный альбумин составляет примерно 60% от общего количества белков плазмы крови. Молекулярный
Сывороточный альбумин составляет примерно 60% от общего количества белков плазмы крови. Молекулярный

Слайд 71 Глобулины — слаборастворимые в воде белки, растворимы в разбавленных растворах солей, входят
Глобулины — слаборастворимые в воде белки, растворимы в разбавленных растворах солей, входят

Слайд 72Гистоны белки, обладающие щелочными свойствами и входящие в состав комплексов с ДНК
Гистоны белки, обладающие щелочными свойствами и входящие в состав комплексов с ДНК

Слайд 73Сложные белки содержат небелковый компонент — простетическую группу.
В зависимости от
Сложные белки содержат небелковый компонент — простетическую группу.
В зависимости от

Слайд 74Металлопротеины белки, в составе которых имеются ионы металлов (медь, железо, цинк, молибден,
Металлопротеины белки, в составе которых имеются ионы металлов (медь, железо, цинк, молибден,

Слайд 75Хромопротеины в качестве простетической группы выступают окрашенные соединения. Типичными хромопротеинами являются белок
Хромопротеины в качестве простетической группы выступают окрашенные соединения. Типичными хромопротеинами являются белок
Слайд 76 Фосфопротены в своем составе содержат остатки фосфорной кислоты, связанные с гидроксильной
Фосфопротены в своем составе содержат остатки фосфорной кислоты, связанные с гидроксильной

Слайд 77 Фосфопротены могут терять фосфатную группу (дефосфорилирование),
могут при определенных условиях быть снова
Фосфопротены могут терять фосфатную группу (дефосфорилирование),
могут при определенных условиях быть снова

Слайд 78 Липопротеины белки, содержащие ковалентно связанные липиды. Эти белки встречаются в составе
Липопротеины белки, содержащие ковалентно связанные липиды. Эти белки встречаются в составе
Слайд 79 Гликопротеины содержат в качестве простетической группы ковалентно связанный углеводный компонент.
Гликопротеины
Гликопротеины содержат в качестве простетической группы ковалентно связанный углеводный компонент.
Гликопротеины

Слайд 81 Гликопротеины широко распространены в природе. Они входят в состав клеточных мембран,
Гликопротеины широко распространены в природе. Они входят в состав клеточных мембран,

Слайд 82Нуклеопротеины - состоят из белков и нуклеиновых кислот, крайние рассматриваются как простетические
Нуклеопротеины - состоят из белков и нуклеиновых кислот, крайние рассматриваются как простетические

Слайд 832. Пространственная организация протеинов
2.1. Типы связей в белковых молекулах
Между аминокислотными остатками в
2. Пространственная организация протеинов
2.1. Типы связей в белковых молекулах
Между аминокислотными остатками в
Слайд 84Ковалентные связи
Ковалентные связи
Слайд 85По своей природе пептидная связь является ковалентной. Ее образование происходит за счет
По своей природе пептидная связь является ковалентной. Ее образование происходит за счет

Слайд 86Название пептида образуется из названия
аминокислотных остатков пептидной цепи
путем замены их
Название пептида образуется из названия
аминокислотных остатков пептидной цепи
путем замены их

Слайд 87Пептидная связь
Пептидная связь
Слайд 88Аминокислоты входящие в состав протеинов, называются аминокислотными остатками. Цепь повторяющихся групп –NH-CH-CO-называется
Аминокислоты входящие в состав протеинов, называются аминокислотными остатками. Цепь повторяющихся групп –NH-CH-CO-называется
Слайд 89В пептидах выделяют особую структуру –
пептидную группу.
Ее образуют атомы
В пептидах выделяют особую структуру –
пептидную группу.
Ее образуют атомы

Слайд 90 Свойства пептидной группы:
Пептидная группа имеет жесткую планарную структуру, т.е. все атомы,
Свойства пептидной группы:
Пептидная группа имеет жесткую планарную структуру, т.е. все атомы,

Слайд 91 Пептидные связи очень прочные, и для их химического гидролиза требуются жесткие условия,
Пептидные связи очень прочные, и для их химического гидролиза требуются жесткие условия,
Слайд 92
Дисульфидная связь
(дисульфидные мостики)
Дисульфидные мостики образуются в молекуле белка между
Дисульфидная связь
(дисульфидные мостики)
Дисульфидные мостики образуются в молекуле белка между
Слайд 93Дисульфидная связь
Дисульфидные мостики могут возникать внутри полипептидной цепи
Дисульфидные мостики могут возникать между
Дисульфидная связь
Дисульфидные мостики могут возникать внутри полипептидной цепи
Дисульфидные мостики могут возникать между
Слайд 94Интересно знать!
С помощью химической завивки волосам можно придать другую форму. Для этого
Интересно знать!
С помощью химической завивки волосам можно придать другую форму. Для этого

Слайд 95Ионные связи
Ионные связи возникают между радикалами аминокислотных остатков, имеющих противоположные заряды, например,
Ионные связи
Ионные связи возникают между радикалами аминокислотных остатков, имеющих противоположные заряды, например,

Слайд 96Гидрофобные взаимодействия
Гидрофобные радикалы аминокислот избегают контактов с водой и поэтому стремятся собраться
Гидрофобные взаимодействия
Гидрофобные радикалы аминокислот избегают контактов с водой и поэтому стремятся собраться
Слайд 97Водородные связи
Водородная связь в молекуле белка образуется
между имеющим частично положительный заряд
Водородные связи
Водородная связь в молекуле белка образуется
между имеющим частично положительный заряд

Слайд 98В белках различают два варианта образования водородных связей:
между пептидными группами;
между
В белках различают два варианта образования водородных связей:
между пептидными группами;
между
Слайд 99Ван-дер-ваальсовы силы
Ван-дер-ваальсовы силы имеют электростатическую природу. Они возникают между разноименными полюсами диполя.
Ван-дер-ваальсовы силы
Ван-дер-ваальсовы силы имеют электростатическую природу. Они возникают между разноименными полюсами диполя.

Слайд 100Рассмотренные выше химические связи принимают участие в формировании структуры белковых молекул
Рассмотренные выше химические связи принимают участие в формировании структуры белковых молекул

Слайд 1012.2. Уровни организации белковых молекул
2.2. Уровни организации белковых молекул
Слайд 1022.2.1. Первичная структура
Первичная структура белка представляет собой линейную цепь аминокислот, расположенных в
2.2.1. Первичная структура
Первичная структура белка представляет собой линейную цепь аминокислот, расположенных в
Слайд 103Первичная структура белка характеризуется:
аминокислотным составом;
последовательностью аминокислотных остатков;
количеством аминокислотных остатков.
Первичная структура белка характеризуется:
аминокислотным составом;
последовательностью аминокислотных остатков;
количеством аминокислотных остатков.

Слайд 104Например, такое заболевание как серповидноклеточная анемия,
связано с изменением формы эритроцита и невозможностью
Например, такое заболевание как серповидноклеточная анемия,
связано с изменением формы эритроцита и невозможностью
Слайд 105Фред Сенгер Фред Сенгер определил аминокислотную последовательность инсулина, продемонстрировав таким способом, что
Фред Сенгер Фред Сенгер определил аминокислотную последовательность инсулина, продемонстрировав таким способом, что

Слайд 1062.2.2. Вторичная структура
Вторичная структура белка образуется в процессе укладки первичной структуры в
2.2.2. Вторичная структура
Вторичная структура белка образуется в процессе укладки первичной структуры в
Слайд 107 Альфа-спираль
Характерна правозакрученная спираль, параметры которой для различных белков одинаковы.
Характеристики α-спирали:
Альфа-спираль
Характерна правозакрученная спираль, параметры которой для различных белков одинаковы.
Характеристики α-спирали:
Слайд 108 Альфа-спираль
Водородные связи, фиксирующие альфа-спираль расположены параллельно оси спирали и многократно
Альфа-спираль
Водородные связи, фиксирующие альфа-спираль расположены параллельно оси спирали и многократно

Слайд 109Некоторые аминокислотные остатки препятствуют
образованию α-спирали:
Например, если в цепи подряд расположено несколько
Некоторые аминокислотные остатки препятствуют
образованию α-спирали:
Например, если в цепи подряд расположено несколько

Слайд 110В остатке пролина при атоме азота, образующем пептидную связь с другой аминокислотой,
В остатке пролина при атоме азота, образующем пептидную связь с другой аминокислотой,

Слайд 111Бета-складчатая структура- или структура складчатого листа фиксирует два или несколько участков полипептидной
Бета-складчатая структура- или структура складчатого листа фиксирует два или несколько участков полипептидной
Слайд 112Цепи могут быть
параллельны или антипараллельны.
Цепи могут быть
параллельны или антипараллельны.

Слайд 113
ПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА
Фиксируют бета-складчатаю структуру водородные
связи между атомами С=О и NH-групп
ПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА
Фиксируют бета-складчатаю структуру водородные
связи между атомами С=О и NH-групп
Слайд 114
ПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА,
состоящая из трех полипептидных цепей
ПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА,
состоящая из трех полипептидных цепей
Слайд 115
АНТИПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА
АНТИПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА
Слайд 116
АНТИПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА, состоящая из трех полипептидных цепей
АНТИПАРАЛЛЕЛЬНАЯ β-СТРУКТУРА, состоящая из трех полипептидных цепей
Слайд 117
ОБРАЗОВАНИЕ β-СТРУКТУРЫ внутри одной полипептидной цепи
ОБРАЗОВАНИЕ β-СТРУКТУРЫ внутри одной полипептидной цепи
Слайд 1182.2.3. Третичная структура
Третичная структура - это трехмерная архитектура полипептидной цепи – особое
2.2.3. Третичная структура
Третичная структура - это трехмерная архитектура полипептидной цепи – особое
Слайд 119 У разных белков третичной структуры различна.
В формировании третичной структуры участвуют дисульфидные
У разных белков третичной структуры различна.
В формировании третичной структуры участвуют дисульфидные

Слайд 120В фибриллярных белках
(например, коллаген, эластин)
молекулы имеют вытянутую форму и обычно формируют
В фибриллярных белках
(например, коллаген, эластин)
молекулы имеют вытянутую форму и обычно формируют

Слайд 121Фибриллярный белок ФИБРОИН – основной компонент натурального шелка и паутины
Фибриллярный белок ФИБРОИН – основной компонент натурального шелка и паутины
Слайд 122В организме млекопитающих коллаген —составляет 25% общего белка. Этот белок имеет необычный
В организме млекопитающих коллаген —составляет 25% общего белка. Этот белок имеет необычный

Слайд 123 В глобулярных белках молекулы имеют форму шара или эллипса. Обычно гидрофобные
В глобулярных белках молекулы имеют форму шара или эллипса. Обычно гидрофобные

Слайд 124Миоглобин – кислород-связывающий белок. Молекула состоит из одной полипептидной цепи, состоящей из
Миоглобин – кислород-связывающий белок. Молекула состоит из одной полипептидной цепи, состоящей из

Слайд 125Схематическое изображение молекулы миоглобина: главная цепь молекулы в виде ленты, боковые группы
Схематическое изображение молекулы миоглобина: главная цепь молекулы в виде ленты, боковые группы
Слайд 126 Можем ли мы определить трехмерную форму белка по его аминокислотной последовательности?
Сейчас
Можем ли мы определить трехмерную форму белка по его аминокислотной последовательности?
Сейчас

Слайд 1272.2.4. Четвертичная структура
Четвертичная структура -
субъединичная структура белка, представляющая взаимное расположение нескольких
2.2.4. Четвертичная структура
Четвертичная структура -
субъединичная структура белка, представляющая взаимное расположение нескольких
Слайд 128Число протомеров может варьировать в широких пределах.
Чаще в молекуле олигомерного белка
Число протомеров может варьировать в широких пределах.
Чаще в молекуле олигомерного белка

Слайд 129Молекула гемоглобина состоит из гема и четырех полипептидных цепей двух различных типов
Молекула гемоглобина состоит из гема и четырех полипептидных цепей двух различных типов

Слайд 130Гем –комплекс Fe (II) с одним из порфиринов – протопорфирином IX. Порфирин
Гем –комплекс Fe (II) с одним из порфиринов – протопорфирином IX. Порфирин
Слайд 131Образование четвертичной структуры глобулярного белка ферритина при объединении молекул в единый ансамбль
Образование четвертичной структуры глобулярного белка ферритина при объединении молекул в единый ансамбль
Слайд 132Многоуровневая организация белковой молекулы
Первичная структура (Последовательность (цепочка) аминокислот)
Вторичная структура (α-спираль, β-слой)
Третичная структура
Многоуровневая организация белковой молекулы
Первичная структура (Последовательность (цепочка) аминокислот)
Вторичная структура (α-спираль, β-слой)
Третичная структура

Слайд 133Многоуровневая организация белковой молекулы
Многоуровневая организация белковой молекулы
Слайд 1342.3. Фолдинг
Правильное сворачивание ( фолдинг ) полипептидных цепей некоторых белков в клетках
2.3. Фолдинг
Правильное сворачивание ( фолдинг ) полипептидных цепей некоторых белков в клетках

Слайд 135Шапероны обеспечивают:
укладку новосинтезированные белков после выхода с рибосом в стабильную трехмерную
Шапероны обеспечивают:
укладку новосинтезированные белков после выхода с рибосом в стабильную трехмерную

Слайд 1363.1. Общие свойства
Характерными физическими свойствами белков являются:
высокая вязкость растворов,
способность к
Характерными физическими свойствами белков являются:
высокая вязкость растворов,
способность к

Слайд 137Молекулярная масса белков находится в пределах 10 000 - 1 000 000.
Молекулярная масса белков находится в пределах 10 000 - 1 000 000.
Слайд 138Сложность состава и высочайшую
молекулярную массу белков можно подтвердить даже этой малой
Сложность состава и высочайшую
молекулярную массу белков можно подтвердить даже этой малой

Слайд 139НАТИВНОСТЬ - это уникальный комплекс физических, физико-химических, химических и биологических свойств белковой
НАТИВНОСТЬ - это уникальный комплекс физических, физико-химических, химических и биологических свойств белковой

Слайд 140Гидролиз - расщеплению полипептидных связей (разрушение первичной структуры) при нагревании белков и
Гидролиз - расщеплению полипептидных связей (разрушение первичной структуры) при нагревании белков и
Слайд 141Аминокислоты, полученные в результате гидролиза, расходуются на синтез собственного белка (увеличение белковой
Аминокислоты, полученные в результате гидролиза, расходуются на синтез собственного белка (увеличение белковой
Слайд 142
3.2.Электрические свойства белковых молекул
Определяются присутствием на поверхности молекул белка положительно и отрицательно
3.2.Электрические свойства белковых молекул
Определяются присутствием на поверхности молекул белка положительно и отрицательно

Слайд 143Значение рН, при котором суммарный заряд белка равен нулю, называется
изоэлектрической точкой
Значение рН, при котором суммарный заряд белка равен нулю, называется
изоэлектрической точкой
Слайд 144 Изоэлектрическая точка для большинства белков находится в области рН от 4,5 до
Изоэлектрическая точка для большинства белков находится в области рН от 4,5 до

Слайд 145В изоэлектрической точке растворимость белка минимальна, так как его молекулы в таком

Слайд 146
3.3.Растворимость белков
Белки бывают растворимые и нерастворимые в воде. Растворимость белков зависит от:
3.3.Растворимость белков
Белки бывают растворимые и нерастворимые в воде. Растворимость белков зависит от:
Слайд 147 Растворимость определяется наличием на поверхности молекул белков заряженных и полярных группировок
Растворимость определяется наличием на поверхности молекул белков заряженных и полярных группировок
Слайд 148Поверхность большинства белковых молекул заряжена отрицательно, и диполи молекул воды притягиваются к
Поверхность большинства белковых молекул заряжена отрицательно, и диполи молекул воды притягиваются к
Слайд 149Чем больше гидрофильных свойств у белковой молекулы, чем больше в ее составе
Чем больше гидрофильных свойств у белковой молекулы, чем больше в ее составе
Слайд 150Реакции осаждения делят на два вида.
Обратимое осаждение белков предполагает выпадение белка в
Реакции осаждения делят на два вида.
Обратимое осаждение белков предполагает выпадение белка в

Слайд 151Обратимое осаждение белков Для данного типа осаждения белков употребляют соли щелочных и
Обратимое осаждение белков Для данного типа осаждения белков употребляют соли щелочных и

Слайд 152Осаждения с потерей нативных свойств белка - процесс необратимый. С белка снимается
Осаждения с потерей нативных свойств белка - процесс необратимый. С белка снимается

Слайд 153При кипячении белок выпадает в осадок. При нагревании до высоких температур усиливается
При кипячении белок выпадает в осадок. При нагревании до высоких температур усиливается

Слайд 154Соли тяжелых металлов образуют соединения с SH-группами белков. Ядовиты для человека и
Соли тяжелых металлов образуют соединения с SH-группами белков. Ядовиты для человека и

Слайд 155Существуют белки, которые растворяются только в растворах солей и не растворяются в

Слайд 156Водорастворимые белки образуют в воде коллоидные растворы.
Эти растворы характеризуются высокой вязкостью, способностью
Эти растворы характеризуются высокой вязкостью, способностью

Слайд 157Явление диализа лежит в основе действия аппарата "искусственная почка", который широко используется
Явление диализа лежит в основе действия аппарата "искусственная почка", который широко используется
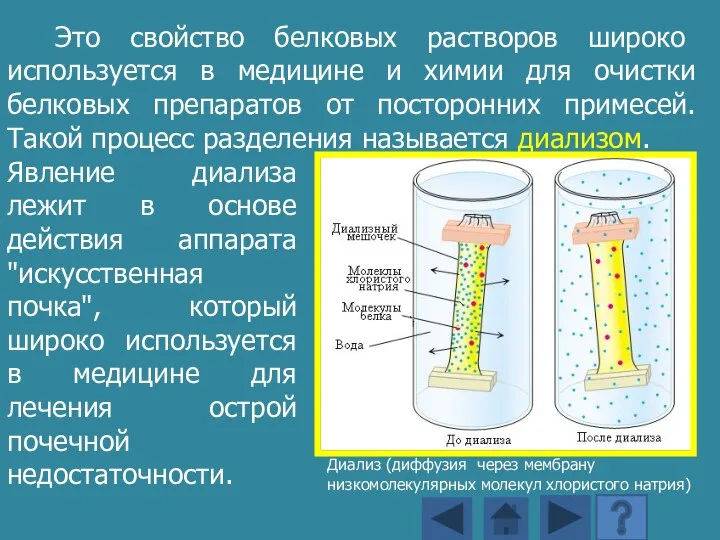
Слайд 158При определенных условиях белки могут образовывать гели. При образовании геля молекулы белка
При определенных условиях белки могут образовывать гели. При образовании геля молекулы белка

Слайд 159 Денатурация - утрата белком природной (нативной) конформации, сопровождающаяся обычно потерей его
Денатурация - утрата белком природной (нативной) конформации, сопровождающаяся обычно потерей его
Слайд 160Факторы, вызывающие денатурацию
Физические факторы
Высокие температуры. Для разных белков характерна различная
Факторы, вызывающие денатурацию
Физические факторы
Высокие температуры. Для разных белков характерна различная

Слайд 161Денатурация может быть обратимой (ренатурация) и необратимой.
Самый известный случай денатурации белка
Денатурация может быть обратимой (ренатурация) и необратимой.
Самый известный случай денатурации белка

Слайд 162Денатурация белков происходит в желудке, где имеется сильнокислая среда (рН 0,5 -

Слайд 163
Ренатурация (обратимая денатурация) – восстановление физико-химических и биологических свойств белка (нативной структуры)
Ренатурация (обратимая денатурация) – восстановление физико-химических и биологических свойств белка (нативной структуры)

Слайд 164При денатурации белков в клетках, в живом организме вырабатываются специфические белки шапероны,
При денатурации белков в клетках, в живом организме вырабатываются специфические белки шапероны,

Слайд 1653.5. Качественные (цветные) реакции на белки
3.5. Качественные (цветные) реакции на белки

Слайд 1663.5. Качественные реакции на белки
Биуретовая реакция – фиолетовое окрашивание при действии солей
3.5. Качественные реакции на белки
Биуретовая реакция – фиолетовое окрашивание при действии солей

Слайд 167Реакция обязана наличию пептидных связей в белках а также продуктах их гидролиза
Реакция обязана наличию пептидных связей в белках а также продуктах их гидролиза

Слайд 168Нингидриновая реакция
Сущность реакции состоит в образовании соединения, окрашенного в сине-фиолетовый цвет,
Нингидриновая реакция
Сущность реакции состоит в образовании соединения, окрашенного в сине-фиолетовый цвет,

Слайд 169При нагревании белка с водным раствором нингидрина аминокислоты окисляются и распадаются, образуя
При нагревании белка с водным раствором нингидрина аминокислоты окисляются и распадаются, образуя
Слайд 170Восстановленный нингидрин конденсируется с аммиаком и окисленной молекулой нингидрина, образуя соединение фиолетово-синего
Восстановленный нингидрин конденсируется с аммиаком и окисленной молекулой нингидрина, образуя соединение фиолетово-синего
Слайд 171Интенсивность окраски полученного сине-фиолетового комплекса (пурпур Руэманна), пропорциональна количеству α-аминокислот.
Желатин
Яичный альбумин
Интенсивность окраски полученного сине-фиолетового комплекса (пурпур Руэманна), пропорциональна количеству α-аминокислот.
Желатин
Яичный альбумин

Слайд 172Ксантопротеиновая реакция – появление желтого окрашивания при действии концентрированной азотной кислоты на
Ксантопротеиновая реакция – появление желтого окрашивания при действии концентрированной азотной кислоты на

Слайд 173 Реакция характерна для некоторых ароматических аминокислот (фенилаланина, тирозина, триптофана), а также
Реакция характерна для некоторых ароматических аминокислот (фенилаланина, тирозина, триптофана), а также
Слайд 174При действии азотной кислоты образуется нитросоединение
желтого цвета.
Нитропроизводные могут реагировать со
При действии азотной кислоты образуется нитросоединение
желтого цвета.
Нитропроизводные могут реагировать со

Слайд 175Реакция Фоля
Реакция на серусодержащие аминокислоты, в ходе которой появляется черное либо
Реакция Фоля
Реакция на серусодержащие аминокислоты, в ходе которой появляется черное либо

Слайд 176 Сульфид натрия Na2S, взаимодействуя с плюмбитом натрия ( появляется в ходе
Сульфид натрия Na2S, взаимодействуя с плюмбитом натрия ( появляется в ходе
Слайд 177Тепловая денатурация белка
При нагревании белки денатурируют. На процесс денатурации оказывают
сильное влияние
Тепловая денатурация белка
При нагревании белки денатурируют. На процесс денатурации оказывают
сильное влияние

Слайд 178Тепловая денатурация белка
В пять пронумерованных пробирок налили по 10 капель раствора яичного
Тепловая денатурация белка
В пять пронумерованных пробирок налили по 10 капель раствора яичного
Слайд 179Тепловая денатурация белка
К раствору белка во второй пробирке добавили одну каплю 1
Тепловая денатурация белка
К раствору белка во второй пробирке добавили одну каплю 1

Слайд 180Тепловая денатурация белка
К раствору белка в третьей пробирке прибавили
1 – 2
Тепловая денатурация белка
К раствору белка в третьей пробирке прибавили
1 – 2

Слайд 181Тепловая денатурация белка
В четвертую пробирку добавили 1 – 2 капли 10 %-го
Тепловая денатурация белка
В четвертую пробирку добавили 1 – 2 капли 10 %-го

Слайд 182Тепловая денатурация белка
В пятую пробирку прибавили 1 – 2 капли насыщенного раствора
Тепловая денатурация белка
В пятую пробирку прибавили 1 – 2 капли насыщенного раствора
Слайд 183Интересно знать!
Прочность белковых молекул просто поразительна! Человеческий волос прочнее меди и может
Интересно знать!
Прочность белковых молекул просто поразительна! Человеческий волос прочнее меди и может

Слайд 184Белки могут выполнять все известные в природе функции, кроме одной - синтезировать
Белки могут выполнять все известные в природе функции, кроме одной - синтезировать

Слайд 1854.1. Ферментативная функция
Самый многообразный и наиболее специализированный класс белков составляют ферменты –
4.1. Ферментативная функция
Самый многообразный и наиболее специализированный класс белков составляют ферменты –

Слайд 186Ферменты
Некоторые из катализируемых реакций, такие как гидролитическое расщепление мочевины, очень просты:
Другие, например
Ферменты
Некоторые из катализируемых реакций, такие как гидролитическое расщепление мочевины, очень просты:
Другие, например
Слайд 187Катализирует преквращение этанола и других алифатических спиртов.
Алкогольдегидрогеназа
Алкогольдегидрогеназа
Слайд 188 У высших животных процесс связывания углекислого газа ускоряется ферментом, который называется
У высших животных процесс связывания углекислого газа ускоряется ферментом, который называется
Слайд 189 В процессе пищеварения участвуют группы ферментов катализирующие расщепление определенных веществ:
– амилазы,
В процессе пищеварения участвуют группы ферментов катализирующие расщепление определенных веществ:
– амилазы,

Слайд 190Интересно знать!
Созданы препараты содержащие ферменты поджелудочной железы.
Например, мезим-форте®, который содержит
Интересно знать!
Созданы препараты содержащие ферменты поджелудочной железы.
Например, мезим-форте®, который содержит

Слайд 191Коллаген
Фибриллярный белок.
Содержится в соединительной ткани: в сухожилиях, костях, хрящах, стенках
Коллаген
Фибриллярный белок.
Содержится в соединительной ткани: в сухожилиях, костях, хрящах, стенках
Слайд 192Интересно знать!
Коллаген является наиболее известным препаратом для коррекции морщин и других дефектов
Интересно знать!
Коллаген является наиболее известным препаратом для коррекции морщин и других дефектов

Слайд 193Кератин
Нерастворимый прочный белок.
Содержится
в волосах, рогах,
копытах, шерсти,
ногтях,
Кератин
Нерастворимый прочный белок.
Содержится
в волосах, рогах,
копытах, шерсти,
ногтях,

Слайд 194Структурные белки
Эластин
Cтруктурный белок способный растягиваться в двух измерениях.
Содержится в связках,
Структурные белки
Эластин
Cтруктурный белок способный растягиваться в двух измерениях.
Содержится в связках,

Слайд 195Фиброин
Главный компонент шелковых нитей и паутины.
Имеет мягкие подвижные
Фиброин
Главный компонент шелковых нитей и паутины.
Имеет мягкие подвижные

Слайд 196Структурные белки
Белки принимают участие в формировании клеточных мембран (в комплексе с фосфолипидами),
Структурные белки
Белки принимают участие в формировании клеточных мембран (в комплексе с фосфолипидами),
Слайд 1974.3. Транспортная функция
Транспортные белки переносят специфические молекулы и ионы.
Внутрь клетки через
4.3. Транспортная функция
Транспортные белки переносят специфические молекулы и ионы.
Внутрь клетки через

Слайд 198Бактериородопсин
проводит протон через мембрану. Сидящие на
a-спиралях бактериородопсина гидрофобные группы обращены
Бактериородопсин
проводит протон через мембрану. Сидящие на
a-спиралях бактериородопсина гидрофобные группы обращены

Слайд 199У многоклеточных организмов существует система транспорта веществ от одних органов к другим.
У многоклеточных организмов существует система транспорта веществ от одних органов к другим.
Слайд 200Интересно знать!
Гемокианин
(от греч.gemo - кровь
и kianos - лазурный,голубой) –кислородпереносящий белок с
Интересно знать!
Гемокианин
(от греч.gemo - кровь
и kianos - лазурный,голубой) –кислородпереносящий белок с
Слайд 201Интересно знать!
Кислород переносится «голубой кровью» у кальмаров, омаров, крабов, осьминогов, каракатиц,
Интересно знать!
Кислород переносится «голубой кровью» у кальмаров, омаров, крабов, осьминогов, каракатиц,

Слайд 202Транспортные белки
В плазме крови многоклеточных организмов находится транспортный белок - сывороточный альбумин,
Транспортные белки
В плазме крови многоклеточных организмов находится транспортный белок - сывороточный альбумин,

Слайд 203В семенах многих растений запасены пищевые белки, потребляемые на первых стадиях развития
В семенах многих растений запасены пищевые белки, потребляемые на первых стадиях развития

Слайд 204К этой группе относятся белки, защищающие организм от вторжения других организмов или
К этой группе относятся белки, защищающие организм от вторжения других организмов или

Слайд 205Защитная реакция наблюдается и при трансплантации. Это грозное явление отторжения чужеродной ткани
Защитная реакция наблюдается и при трансплантации. Это грозное явление отторжения чужеродной ткани
Слайд 206Фибрин - нерастворимый белок, образующийся в процессе свертывания крови. Нити фибрина, полимеризуясь,
Фибрин - нерастворимый белок, образующийся в процессе свертывания крови. Нити фибрина, полимеризуясь,
Слайд 207Стрессовые белки синтезируются в растениях в ответ на различные экстремальные воздействия: окислительный
Стрессовые белки синтезируются в растениях в ответ на различные экстремальные воздействия: окислительный

Слайд 208 Схема ответа белков теплового шока (БТШ) в растительных клетках: зеленые волнистые
Схема ответа белков теплового шока (БТШ) в растительных клетках: зеленые волнистые
Слайд 209Белки - рецепторы
«Благоденствие» и выживание каждой клетки и организма в целом зависят
Белки - рецепторы
«Благоденствие» и выживание каждой клетки и организма в целом зависят
Слайд 210Белки - рецепторы
Существует три основных типа рецепторов,
интегрированных во внешнюю клеточную мембрану:
рецепторы,
Белки - рецепторы
Существует три основных типа рецепторов,
интегрированных во внешнюю клеточную мембрану:
рецепторы,

Слайд 211Рецепторы, сопряженные с G–белками
G- белки – это регуляторы, включающие или выключающие
Рецепторы, сопряженные с G–белками
G- белки – это регуляторы, включающие или выключающие

Слайд 212Структура холинэргического рецептора никотинового типа, формирующего ионный канал. Субъединицы полипептидной цепи пронизывают
Структура холинэргического рецептора никотинового типа, формирующего ионный канал. Субъединицы полипептидной цепи пронизывают
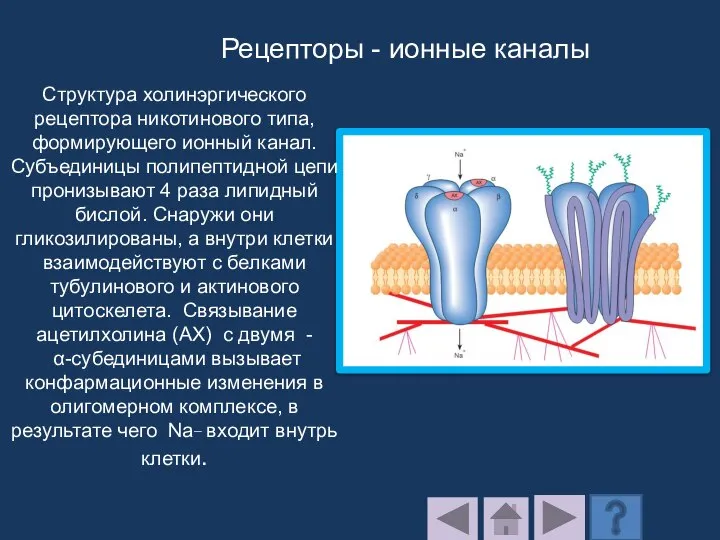
Слайд 213Рецепторы, ассоциированные с ферментативной активностью
Белки-рецепторы данного типа могут
обладать собственной активностью,
Рецепторы, ассоциированные с ферментативной активностью
Белки-рецепторы данного типа могут
обладать собственной активностью,
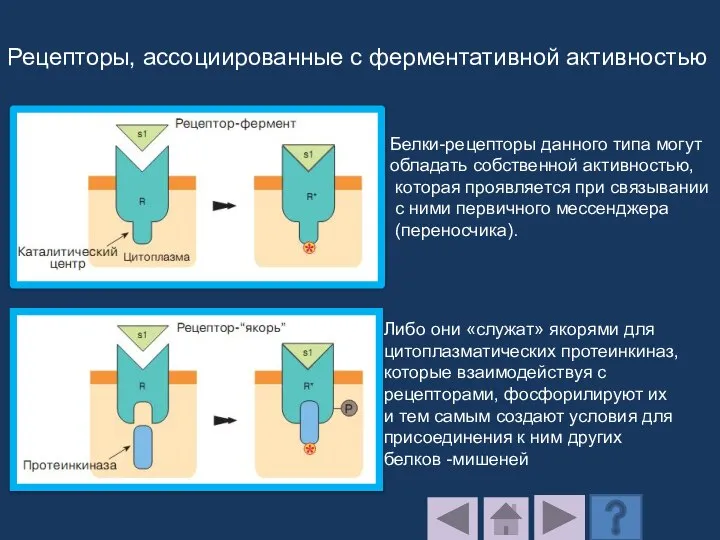
Слайд 214Некоторые белки наделяют клетку или организм способностью сокращаться, изменять форму или передвигаться.
Некоторые белки наделяют клетку или организм способностью сокращаться, изменять форму или передвигаться.
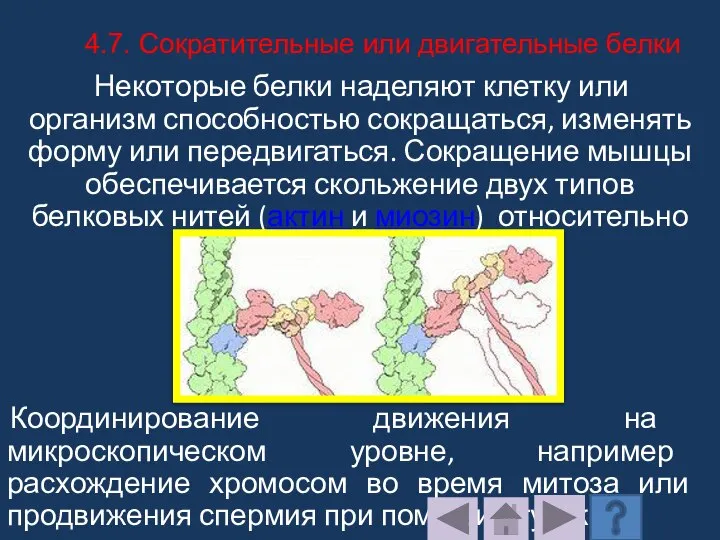
Слайд 215Белки обладают чрезвычайно широкой биологической активностью. Они регулируют работу сердечно-сосудистой, иммунной, половой,
Белки обладают чрезвычайно широкой биологической активностью. Они регулируют работу сердечно-сосудистой, иммунной, половой,

Слайд 216К ним принадлежат некоторые гормоны, вырабатываемые в железах внутренней секреции. Инсулин и
К ним принадлежат некоторые гормоны, вырабатываемые в железах внутренней секреции. Инсулин и
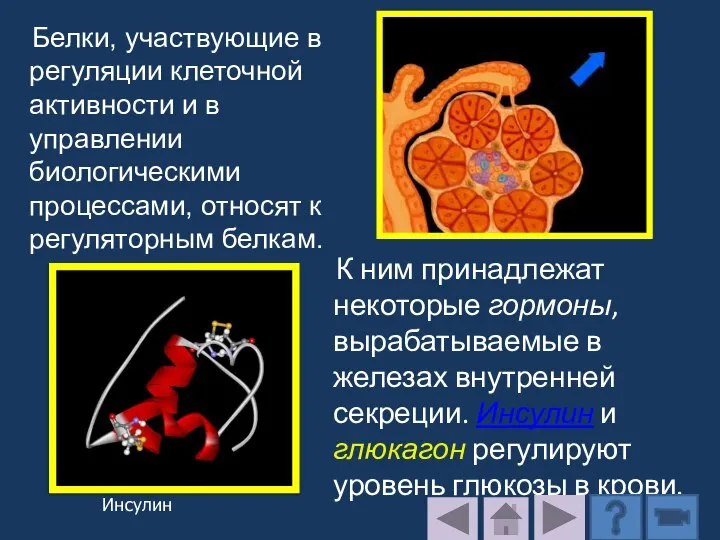
Слайд 217Окситоцин человека
Пептидные гормоны
Окситоцин
Стимулирует сокращение матки у беременных самок.
Стимулирует выделение
Окситоцин человека
Пептидные гормоны
Окситоцин
Стимулирует сокращение матки у беременных самок.
Стимулирует выделение

Слайд 218Вазопрессин человека
Пептидные гормоны
Вазопрессин
Обладает сильным антидиуретическим действием (регулирует водный обмен), стимулируя
Вазопрессин человека
Пептидные гормоны
Вазопрессин
Обладает сильным антидиуретическим действием (регулирует водный обмен), стимулируя

Слайд 219Гормон роста (соматотропин) определяет размеры тела, секретируется передней долей гипофиза. Гормоном роста
Гормон роста (соматотропин) определяет размеры тела, секретируется передней долей гипофиза. Гормоном роста

Слайд 220Недостаток гормона роста в детском возрасте связан в основном с генетическими дефектами
Недостаток гормона роста в детском возрасте связан в основном с генетическими дефектами

Слайд 2214.9. Токсины
Большая группа фармакологически активных пептидов, в том числе брадикинин, физалемин, церулеин
4.9. Токсины
Большая группа фармакологически активных пептидов, в том числе брадикинин, физалемин, церулеин

Слайд 222Некоторые из них вызывают быстрое сокращение внесосудистой мускулатуры и получили название тахикининов.
Брадикинин
Некоторые из них вызывают быстрое сокращение внесосудистой мускулатуры и получили название тахикининов. Брадикинин
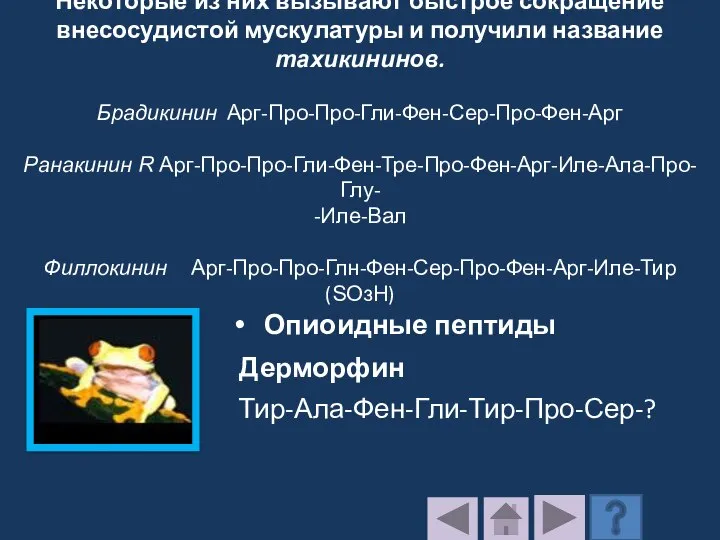
Слайд 223Токсины
Мелиттин — основной компонент пчелиного яда, содержание его в целом яде
Токсины
Мелиттин — основной компонент пчелиного яда, содержание его в целом яде

Слайд 224токсины
Все токсические полипептиды змей делят на:
- токсины, специфически блокирующие Н-холинорецепторы субсинаптической мембраны
токсины
Все токсические полипептиды змей делят на:
- токсины, специфически блокирующие Н-холинорецепторы субсинаптической мембраны

Слайд 225токсины
Бледная поганка
Смертельно ядовитый гриб.
α-аманитин
токсины
Бледная поганка
Смертельно ядовитый гриб.
α-аманитин
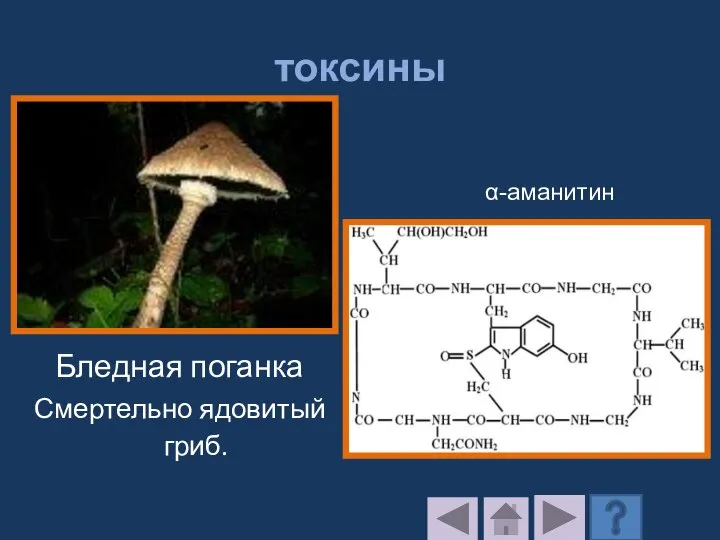
Слайд 226 Интересно знать!
Нобелевская премия по химии 2008 года присуждена трём биологам —
Интересно знать!
Нобелевская премия по химии 2008 года присуждена трём биологам —

Слайд 227Белки используют
в промышленности
для окрашивания тканей,
при рафинировании сахара,
в различных
Белки используют
в промышленности
для окрашивания тканей,
при рафинировании сахара,
в различных
Слайд 228Интересно знать!
В плазме некоторых антарктических рыб содержатся белки со свойствами антифриза,
Интересно знать!
В плазме некоторых антарктических рыб содержатся белки со свойствами антифриза,

Слайд 229Кроме выше рассмотренных функций, белки участвуют в экспрессии генетической информации, передаче нервных
Кроме выше рассмотренных функций, белки участвуют в экспрессии генетической информации, передаче нервных

Слайд 230Белки как необходимые компоненты пищи
Необходимые для человека белки содержат продукты питания
Белки как необходимые компоненты пищи
Необходимые для человека белки содержат продукты питания

Слайд 231Содержание белка в животных продуктах
Мясо, рыба и птица усваиваются организмом в разной
Содержание белка в животных продуктах
Мясо, рыба и птица усваиваются организмом в разной

Слайд 232Содержание белка в растительных продуктах
Бобовые и крупы отличаются особенно высоким содержанием белка.
Содержание белка в растительных продуктах
Бобовые и крупы отличаются особенно высоким содержанием белка.

Слайд 233Дефицит белка в питании уменьшает устойчивость организма к инфекциям, так как снижается
Дефицит белка в питании уменьшает устойчивость организма к инфекциям, так как снижается

Слайд 234Заключение
Сегодня на «коротком» пути от гена к белку (он весь умещается в
Заключение
Сегодня на «коротком» пути от гена к белку (он весь умещается в

Слайд 235После того, как синтезирована аминокислотная последовательность белка, получившийся продукт еще не стал
После того, как синтезирована аминокислотная последовательность белка, получившийся продукт еще не стал

Слайд 236Пространственную структуру белка сегодня можно определить лишь экспериментально - с помощью рентгеноструктурного
Пространственную структуру белка сегодня можно определить лишь экспериментально - с помощью рентгеноструктурного

 Профессии, связанне с химией
Профессии, связанне с химией Кислоты. Химический диктант. 8 класс
Кислоты. Химический диктант. 8 класс Основы молекулярно-кинетической теории
Основы молекулярно-кинетической теории Применение сверхкритических флюидных технологий в химии природных соединений
Применение сверхкритических флюидных технологий в химии природных соединений Study of short peptide adsorption on solution dispersed. Inorganic nanoparticles using depletion method
Study of short peptide adsorption on solution dispersed. Inorganic nanoparticles using depletion method Минералы. Самородные элементы
Минералы. Самородные элементы Водные растворы электролитов
Водные растворы электролитов Галогены. 9 класс
Галогены. 9 класс Melatonin. Краткий экскурс
Melatonin. Краткий экскурс Химическая связь в твердых телах. Экзаменационные вопросы
Химическая связь в твердых телах. Экзаменационные вопросы Классы неорганических соединений (основания)
Классы неорганических соединений (основания) Презентация на тему Алкадиены: строение, номенклатура, гомологи, изомерия
Презентация на тему Алкадиены: строение, номенклатура, гомологи, изомерия  електроліти та неелектроліти
електроліти та неелектроліти Жидкая углекислота
Жидкая углекислота Свойства воды
Свойства воды Этиленовые углеводороды
Этиленовые углеводороды Альдегиды (номенклатура и изомерия)
Альдегиды (номенклатура и изомерия) Homecredit Bank. Показатели и зоны роста
Homecredit Bank. Показатели и зоны роста Мило. Його склад. Мийна дія
Мило. Його склад. Мийна дія Получение этилового спирта из древесины
Получение этилового спирта из древесины Гидролиз солей
Гидролиз солей Основные диагностические признаки рубина и его имитаций
Основные диагностические признаки рубина и его имитаций Химия и искусство: техника офорт
Химия и искусство: техника офорт Закон Гесса. Термохимические уравнения
Закон Гесса. Термохимические уравнения Хлор. Химические свойства и применение. Лекция 5
Хлор. Химические свойства и применение. Лекция 5 Химические вещества парабены
Химические вещества парабены Железо как химический элемент
Железо как химический элемент Применение кислорода
Применение кислорода