- Общая физиология центральной нервной системы. Рефлекторный принцип нервной регуляции

Содержание
- 2. Физиология центральной нервной системы (ЦНС) является наиболее сложной, но в то же время и наиболее ответственной
- 3. Рис. 9. Схема рефлекторного кольца Ученик Павлова П.К. Анохин, создав концепцию о функциональной системе, как основном
- 4. Такая структура рефлекторной дуги превращает ее в самонастраивающийся нервный контур регуляции физиологической функции, совершенствуя рефлекторную реакцию
- 5. 5-2. НЕЙРОНЫ ЦНС: КЛАССИФИКАЦИЯ, СВОЙСТВА НЕРВНЫХ КЛЕТОК. ВИДЫ СВЯЗЕЙ МЕЖДУ НЕЙРОНАМИ В ЦНС. Структурной и функциональной
- 6. Рис. 10. Строение нейрона и электронно-микроскопическая картина строения тела клетки Структура и размеры нейронов сильно варьируют.
- 7. Место отхождения аксона от тела нервной клетки (аксонный холмик) имеет наибольшее значение в возбуждении нейрона. Это
- 8. Особенности метаболизма нейронов. 1. В нервных клетках потребление кислорода и глюкозы выше, чем у других клеток
- 9. Классификация нейронов Нейроны в ЦНС разделяют на афферентные (чувствительные), эфферентные (эффекторные) и промежуточные, или вставочные (ассоциативные).
- 10. Нервные клетки обычно окружены вспомогательными глиальными клетками, занимающими примерно 50% объема ЦНС. Их число превышает число
- 11. Нейронные сети Механизмы мозга, в частности реализация психических функций, определяются свойствами нейрона как элемента нейронной сети.
- 12. 5.3. КЛАССИФИКАЦИЯ СИНАПСОВ ЦНС, МЕДИАТОРЫ СИНАПСОВ ЦНС И ИХ ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ. СВОЙСТВА СИНАПСОВ ЦНС. Синапсами называются
- 13. Для синапсов с химическим способом передачи возбуждения характерны синаптическая задержка проведения возбуждения, длящаяся около 0,5 мс,
- 14. В пресинаптической терминали образуется и скапливается в виде пузырьков медиатор, например, ацетилхолин. В каждом пузырьке находится
- 15. Этапы синаптической передачи. Синаптическая передача возможна при реализации ряда последовательных процессов: синтез нейромедиатора, его накопление и
- 16. 6. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны либо прямо (поступление ионов
- 17. Характеристика отдельных нейромедиаторов Ацетилхолин секретируется из терминалей соматических мотонейронов (нервно-мышечные синапсы), преганглионарных волокон, постганглионарных холинергических (парасимпатических)
- 18. Серотонин (5-гидрокситриптамин) — нейромедиатор многих центральных нейронов (например, ядра шва, нейроны восходящей ретикулярной активирующей системы). Предшественником
- 19. Динорфины. Эта группа нейромедиаторов состоит из 7 пептидов близкой аминокислотной последовательности, которые присутствуют в нейронах тех
- 20. Свойства синапсов в ЦНС 1) Относительная медиаторная специфичность синапса, т. е. каждый синапс имеет свой доминирующий
- 22. Скачать презентацию

Слайд 3Рис. 9. Схема рефлекторного кольца
Ученик Павлова П.К. Анохин, создав концепцию о
Рис. 9. Схема рефлекторного кольца
Ученик Павлова П.К. Анохин, создав концепцию о

Слайд 4Такая структура рефлекторной дуги превращает ее в самонастраивающийся нервный контур регуляции физиологической
Такая структура рефлекторной дуги превращает ее в самонастраивающийся нервный контур регуляции физиологической

Слайд 5 5-2. НЕЙРОНЫ ЦНС: КЛАССИФИКАЦИЯ, СВОЙСТВА НЕРВНЫХ КЛЕТОК. ВИДЫ СВЯЗЕЙ МЕЖДУ НЕЙРОНАМИ
5-2. НЕЙРОНЫ ЦНС: КЛАССИФИКАЦИЯ, СВОЙСТВА НЕРВНЫХ КЛЕТОК. ВИДЫ СВЯЗЕЙ МЕЖДУ НЕЙРОНАМИ

Слайд 6Рис. 10. Строение нейрона и электронно-микроскопическая картина строения тела клетки
Структура и
Рис. 10. Строение нейрона и электронно-микроскопическая картина строения тела клетки
Структура и

Слайд 7Место отхождения аксона от тела нервной клетки (аксонный холмик) имеет наибольшее значение
Место отхождения аксона от тела нервной клетки (аксонный холмик) имеет наибольшее значение

Слайд 8Особенности метаболизма нейронов.
1. В нервных клетках потребление кислорода и глюкозы выше,
Особенности метаболизма нейронов.
1. В нервных клетках потребление кислорода и глюкозы выше,

Слайд 9Классификация нейронов
Нейроны в ЦНС разделяют на афферентные (чувствительные), эфферентные (эффекторные) и
Классификация нейронов
Нейроны в ЦНС разделяют на афферентные (чувствительные), эфферентные (эффекторные) и

Слайд 10Нервные клетки обычно окружены вспомогательными глиальными клетками, занимающими примерно 50% объема ЦНС.
Нервные клетки обычно окружены вспомогательными глиальными клетками, занимающими примерно 50% объема ЦНС.

Слайд 11Нейронные сети
Механизмы мозга, в частности реализация психических функций, определяются свойствами нейрона как
Нейронные сети
Механизмы мозга, в частности реализация психических функций, определяются свойствами нейрона как

Слайд 125.3. КЛАССИФИКАЦИЯ СИНАПСОВ ЦНС, МЕДИАТОРЫ СИНАПСОВ ЦНС И ИХ ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ. СВОЙСТВА
5.3. КЛАССИФИКАЦИЯ СИНАПСОВ ЦНС, МЕДИАТОРЫ СИНАПСОВ ЦНС И ИХ ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ. СВОЙСТВА

Слайд 13Для синапсов с химическим способом передачи возбуждения характерны синаптическая задержка проведения возбуждения,
Для синапсов с химическим способом передачи возбуждения характерны синаптическая задержка проведения возбуждения,

Слайд 14В пресинаптической терминали образуется и скапливается в виде пузырьков медиатор, например, ацетилхолин.
В пресинаптической терминали образуется и скапливается в виде пузырьков медиатор, например, ацетилхолин.

Слайд 15Этапы синаптической передачи. Синаптическая передача возможна при реализации ряда последовательных процессов: синтез
Этапы синаптической передачи. Синаптическая передача возможна при реализации ряда последовательных процессов: синтез
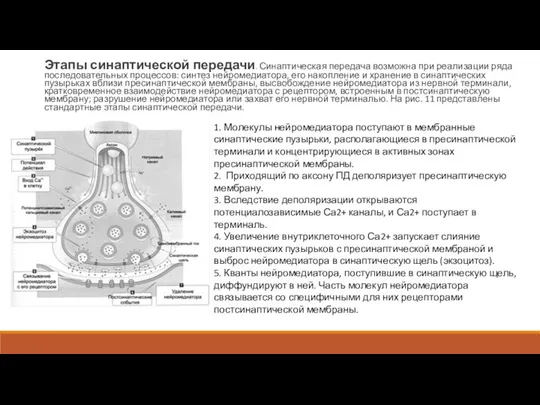
Слайд 166. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны
6. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны

Слайд 17Характеристика отдельных нейромедиаторов
Ацетилхолин секретируется из терминалей соматических мотонейронов (нервно-мышечные синапсы), преганглионарных волокон,
Характеристика отдельных нейромедиаторов
Ацетилхолин секретируется из терминалей соматических мотонейронов (нервно-мышечные синапсы), преганглионарных волокон,

Слайд 18Серотонин (5-гидрокситриптамин) — нейромедиатор многих центральных нейронов (например, ядра шва, нейроны восходящей
Серотонин (5-гидрокситриптамин) — нейромедиатор многих центральных нейронов (например, ядра шва, нейроны восходящей

Слайд 19Динорфины. Эта группа нейромедиаторов состоит из 7 пептидов близкой аминокислотной последовательности, которые
Динорфины. Эта группа нейромедиаторов состоит из 7 пептидов близкой аминокислотной последовательности, которые

Слайд 20Свойства синапсов в ЦНС
1) Относительная медиаторная специфичность синапса, т. е. каждый синапс
Свойства синапсов в ЦНС
1) Относительная медиаторная специфичность синапса, т. е. каждый синапс

 Проблемы распределения донорских органов. Лист ожидания, особенности его оформления. Медицинские и этические критерии
Проблемы распределения донорских органов. Лист ожидания, особенности его оформления. Медицинские и этические критерии Ишемический колит
Ишемический колит Анальная трещина: принципы диагностики и лечения
Анальная трещина: принципы диагностики и лечения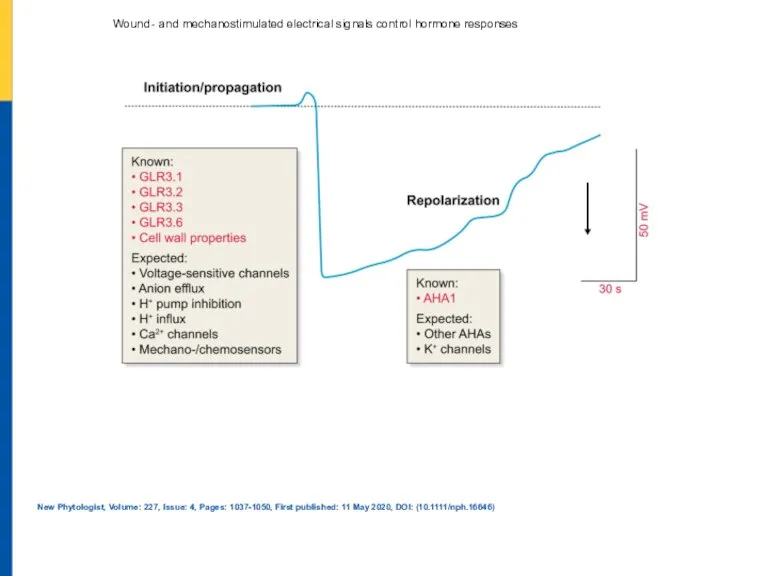 Wound‐ and mechanostimulated electrical signals control hormone responses
Wound‐ and mechanostimulated electrical signals control hormone responses Беременность и роды при инфекциях
Беременность и роды при инфекциях Причины нарушений речи у детей
Причины нарушений речи у детей Всероссийская акция Стоп ВИЧ/СПИД
Всероссийская акция Стоп ВИЧ/СПИД Характеристика трансплантатов, необходимых для пластики передней крестообразной связки
Характеристика трансплантатов, необходимых для пластики передней крестообразной связки Ас қорыту - фармакология
Ас қорыту - фармакология Нарушение тромбоцитарно-сосудистого гемостаза
Нарушение тромбоцитарно-сосудистого гемостаза Оказание услуг общего ухода и помощи при осуществлении повседневной деятельности. Лекция 2
Оказание услуг общего ухода и помощи при осуществлении повседневной деятельности. Лекция 2 Жидкостные компартменты
Жидкостные компартменты Алгоритм по применению грелки
Алгоритм по применению грелки Техника эффективной методики хирургического лечения глаукомы
Техника эффективной методики хирургического лечения глаукомы Инфекции у детей - что происходит сейчас? Туберкулез. Гепатит В
Инфекции у детей - что происходит сейчас? Туберкулез. Гепатит В Осложнения острого аппендицита
Осложнения острого аппендицита Клинические исследования и доказательная медицина
Клинические исследования и доказательная медицина Tutorium Histologie für beruflich
Tutorium Histologie für beruflich Неврологические симптомы и синдромы
Неврологические симптомы и синдромы Что нужно делать, чтобы не заболеть
Что нужно делать, чтобы не заболеть Денсаулық технологиясы
Денсаулық технологиясы Холиномиметики. Классификация холиномиметиков. Препараты
Холиномиметики. Классификация холиномиметиков. Препараты Планирование здравоохранения
Планирование здравоохранения Правила работы с медицинским оборудованием в стоматологии
Правила работы с медицинским оборудованием в стоматологии Противосифилитические средства
Противосифилитические средства Инфекции передаваемые половым путём
Инфекции передаваемые половым путём Хронический панкреатит
Хронический панкреатит Теоретические основы судебной ветеринарно-санитарной экспертизы
Теоретические основы судебной ветеринарно-санитарной экспертизы