- Рецепторы лекарственных веществ, сопряженные с G-белками

Содержание
- 2. G-белок-связанный рецептор представляют собой белки, интегрированные в плазматическую мембрану и состоящие из связки семи гидрофобных спиралей,
- 4. К наиболее распространённым клеточным рецепторам этого типа относят вазопрессиновые и ангиотензиновые, α-адренорецепторы, β-адренорецепторы и м-холинорецепторы, опиатные
- 5. Аденилатциклазный механизм передачи сигнала
- 6. Антагонисты рецепторов ангиотензина II, или блокаторы АТ1-рецепторов — одна из новых групп антигипертензивных средств. Она объединяет
- 7. Для адренорецепторов характерна разная чувствительность к химическим соединениям, и поэтому их делят на α (α1, α2)
- 8. Препараты, стимулирующие адренорецепторы, называют адреномиметиками, а угнетающие – адреноблокаторами (адренолитиками) Адреномиметики По преимущественному влиянию на α-
- 9. βγ-Адреномиметики (стимуляторы βγ-адренорецепторов) β1-Адренорецепторы преимущественно локализованы в сердце в мембране кардиомиоцитов. Стимуляция β1-aдренорецепторов (связанных с Gs-белками)
- 10. М-холиномиметики стимулируют М-холинорецепторы, расположенные в мембране клеток эффекторных органов и тканей, получающих парасимпатическую иннервацию. М-холинорецепторы подразделяют
- 11. М3-холинорецепторы гладкомышечных клеток и клеток экзокринных желез взаимодействуют с Gq-белками, которые активируют фосфолипазу С. При участии
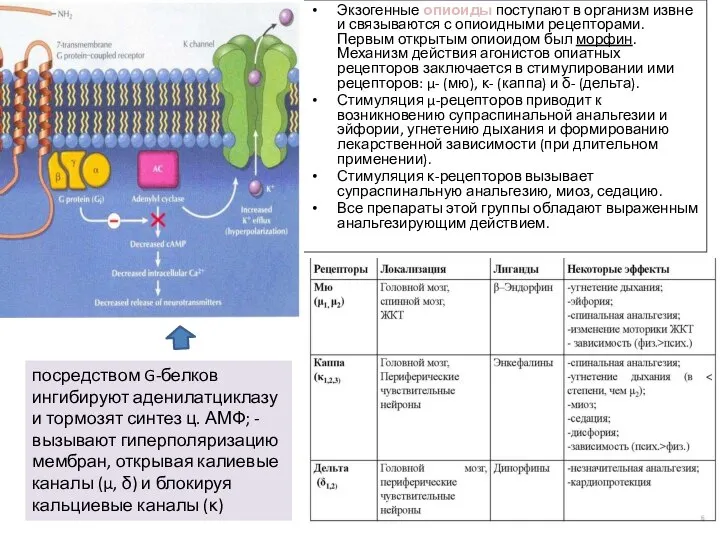
- 13. Экзогенные опиоиды поступают в организм извне и связываются с опиоидными рецепторами. Первым открытым опиоидом был морфин.
- 14. Дофаминовые рецепторы присутствуют как в центральной нервной системе, так в периферических органах. Дофаминомиметики. 1) непрямого действия,
- 15. Дофаминолитики. Препараты нейролептического действия, блокирующие постсинаптические дофаминовые рецепторы с одновременным блокированием пресинаптических дофаминовых рецепторов. Считается, что
- 17. Тест. 1.Что из себя представляет G-белок-связанный рецептор? А) представляют собой белки, интегрированные в плазматическую мембрану и
- 18. Тест. 1.Что из себя представляет G-белок-связанный рецептор? А) представляют собой белки, интегрированные в плазматическую мембрану и
- 19. 2. К G-белок-связанный рецепторам типа не относят: А) ангиотензиновые Б)α-адренорецепторы В)β-адренорецепторы Г)н-холинорецепторы Д) опиатные Е) дофаминовые
- 20. 2. К G-белок-связанный рецепторам типа не относят: А) ангиотензиновые Б)α-адренорецепторы В)β-адренорецепторы Г)н-холинорецепторы ( м-холинорецепторы!) Д) опиатные
- 21. 3. Ацеклидин и пилокарпин — вызывают локальные (при местном применение) или общие эффекты возбуждения ? рецепторов:
- 23. Скачать презентацию
Слайд 2G-белок-связанный рецептор представляют собой белки, интегрированные в плазматическую мембрану и состоящие из
G-белок-связанный рецептор представляют собой белки, интегрированные в плазматическую мембрану и состоящие из
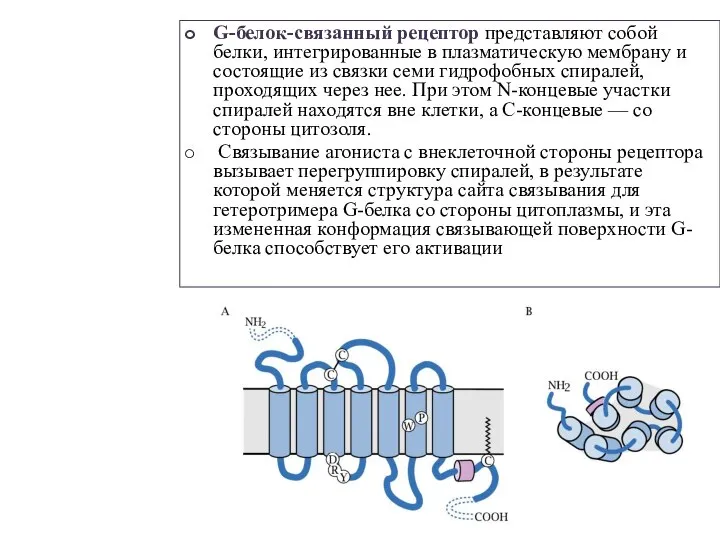
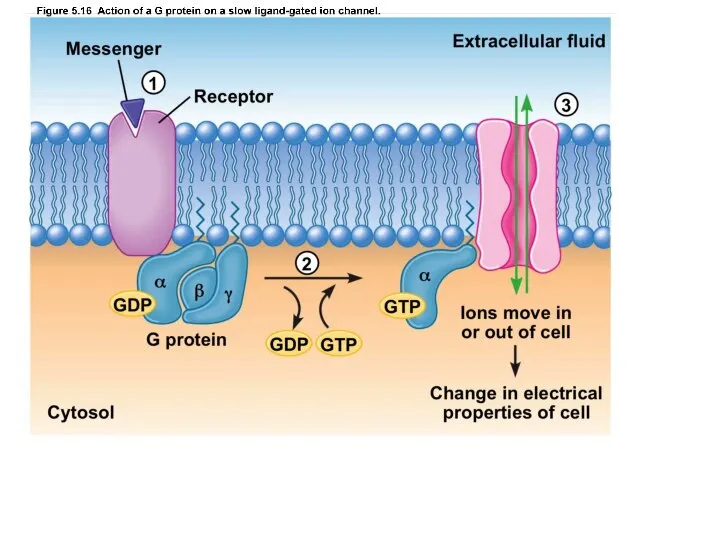
Слайд 4К наиболее распространённым клеточным рецепторам этого типа относят вазопрессиновые и ангиотензиновые, α-адренорецепторы,
К наиболее распространённым клеточным рецепторам этого типа относят вазопрессиновые и ангиотензиновые, α-адренорецепторы,

Слайд 5Аденилатциклазный механизм передачи сигнала
Аденилатциклазный механизм передачи сигнала
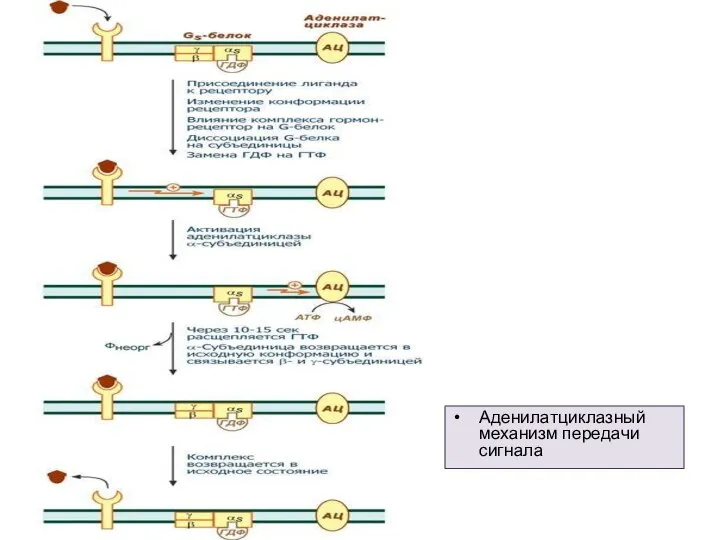
Слайд 6Антагонисты рецепторов ангиотензина II, или блокаторы АТ1-рецепторов — одна из новых групп антигипертензивных
Антагонисты рецепторов ангиотензина II, или блокаторы АТ1-рецепторов — одна из новых групп антигипертензивных

Слайд 7Для адренорецепторов характерна разная чувствительность к химическим соединениям, и поэтому их делят
Для адренорецепторов характерна разная чувствительность к химическим соединениям, и поэтому их делят

Слайд 8Препараты, стимулирующие адренорецепторы, называют адреномиметиками, а угнетающие – адреноблокаторами (адренолитиками)
Адреномиметики
По преимущественному влиянию
Препараты, стимулирующие адренорецепторы, называют адреномиметиками, а угнетающие – адреноблокаторами (адренолитиками)
Адреномиметики
По преимущественному влиянию

Слайд 9βγ-Адреномиметики (стимуляторы βγ-адренорецепторов)
β1-Адренорецепторы преимущественно локализованы в сердце в мембране кардиомиоцитов. Стимуляция β1-aдренорецепторов
βγ-Адреномиметики (стимуляторы βγ-адренорецепторов)
β1-Адренорецепторы преимущественно локализованы в сердце в мембране кардиомиоцитов. Стимуляция β1-aдренорецепторов

Слайд 10М-холиномиметики стимулируют М-холинорецепторы, расположенные в мембране клеток эффекторных органов и тканей, получающих
М-холиномиметики стимулируют М-холинорецепторы, расположенные в мембране клеток эффекторных органов и тканей, получающих

Слайд 11М3-холинорецепторы гладкомышечных клеток и клеток экзокринных желез взаимодействуют с Gq-белками, которые активируют
М3-холинорецепторы гладкомышечных клеток и клеток экзокринных желез взаимодействуют с Gq-белками, которые активируют

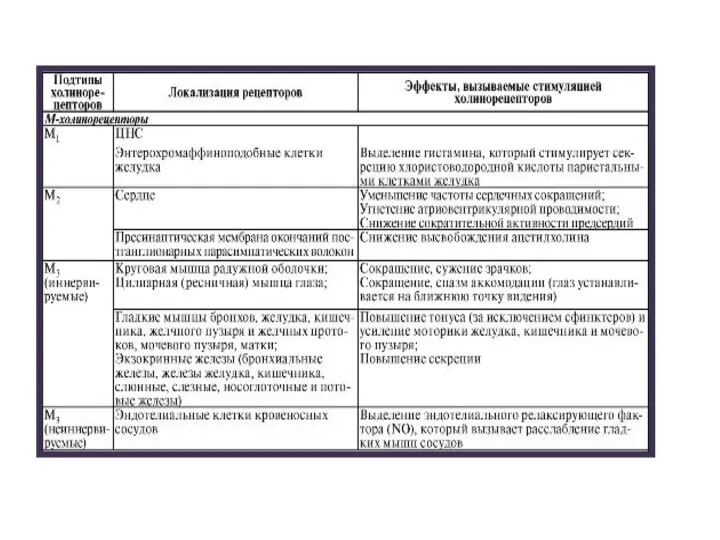
Слайд 13Экзогенные опиоиды поступают в организм извне и связываются с опиоидными рецепторами. Первым
Экзогенные опиоиды поступают в организм извне и связываются с опиоидными рецепторами. Первым
Слайд 14Дофаминовые рецепторы присутствуют как в центральной нервной системе, так в периферических органах.
Дофаминомиметики.
1) непрямого действия,
Дофаминовые рецепторы присутствуют как в центральной нервной системе, так в периферических органах.
Дофаминомиметики.
1) непрямого действия,

Слайд 15Дофаминолитики.
Препараты нейролептического действия, блокирующие постсинаптические дофаминовые рецепторы с одновременным блокированием пресинаптических дофаминовых
Дофаминолитики.
Препараты нейролептического действия, блокирующие постсинаптические дофаминовые рецепторы с одновременным блокированием пресинаптических дофаминовых


Слайд 17Тест.
1.Что из себя представляет G-белок-связанный рецептор?
А) представляют собой белки, интегрированные в плазматическую
Тест.
1.Что из себя представляет G-белок-связанный рецептор?
А) представляют собой белки, интегрированные в плазматическую

Слайд 18Тест.
1.Что из себя представляет G-белок-связанный рецептор?
А) представляют собой белки, интегрированные в плазматическую
Тест.
1.Что из себя представляет G-белок-связанный рецептор?
А) представляют собой белки, интегрированные в плазматическую

Слайд 192. К G-белок-связанный рецепторам типа не относят:
А) ангиотензиновые
Б)α-адренорецепторы
В)β-адренорецепторы
Г)н-холинорецепторы
Д) опиатные
Е) дофаминовые
2. К G-белок-связанный рецепторам типа не относят:
А) ангиотензиновые
Б)α-адренорецепторы
В)β-адренорецепторы
Г)н-холинорецепторы
Д) опиатные
Е) дофаминовые

Слайд 202. К G-белок-связанный рецепторам типа не относят:
А) ангиотензиновые
Б)α-адренорецепторы
В)β-адренорецепторы
Г)н-холинорецепторы ( м-холинорецепторы!)
Д) опиатные
Е)
2. К G-белок-связанный рецепторам типа не относят:
А) ангиотензиновые
Б)α-адренорецепторы
В)β-адренорецепторы
Г)н-холинорецепторы ( м-холинорецепторы!)
Д) опиатные
Е)

Слайд 213. Ацеклидин и пилокарпин — вызывают локальные (при местном применение) или общие
3. Ацеклидин и пилокарпин — вызывают локальные (при местном применение) или общие

 Кровяные инфекции. Чума
Кровяные инфекции. Чума Пролапс митрального клапана и дизъюнкция. Оценка рисков ВСС
Пролапс митрального клапана и дизъюнкция. Оценка рисков ВСС Основы трансфузиологии
Основы трансфузиологии Биогенные амины. Ингибиторы МАО. Антигистаминные препараты
Биогенные амины. Ингибиторы МАО. Антигистаминные препараты МРТ диагностика компрессионной миелопатии, изменений интра - и эпидуральных пространств
МРТ диагностика компрессионной миелопатии, изменений интра - и эпидуральных пространств Токсикоз с эксикозом при кишечных инфекциях у детей
Токсикоз с эксикозом при кишечных инфекциях у детей Уреаплазмоз. Микоплазмоз
Уреаплазмоз. Микоплазмоз Врожденный сифилис. Лекция 7
Врожденный сифилис. Лекция 7 Дневник развития ребенка
Дневник развития ребенка ОВИН – диагноз исключения. Клинические примеры
ОВИН – диагноз исключения. Клинические примеры Сыроедение
Сыроедение Розвиток професійних хвороб зумовлений впливом на організм працівників шкідливих виробничих чинників
Розвиток професійних хвороб зумовлений впливом на організм працівників шкідливих виробничих чинників Глаукомы. Лекция 10
Глаукомы. Лекция 10 Периферический увеит
Периферический увеит Жеке даму биологиясы әдістері
Жеке даму биологиясы әдістері Избыточная масса тела. Ожирение
Избыточная масса тела. Ожирение Опухоли головного мозга
Опухоли головного мозга Паховая грыжа
Паховая грыжа Алкогольная полиневропатия
Алкогольная полиневропатия Простагландины. Функции, используемые для терапии
Простагландины. Функции, используемые для терапии Психофармакотерапия. Анксиолитики. Гипнотики
Психофармакотерапия. Анксиолитики. Гипнотики Роль гиперчувствительности замедленного типа в инфекционном процессе и нестерильном иммунитете
Роль гиперчувствительности замедленного типа в инфекционном процессе и нестерильном иммунитете О речевом развитии детей. Что должны знать родители
О речевом развитии детей. Что должны знать родители Эндокринология. Феохромоцитома
Эндокринология. Феохромоцитома Острая почечная недостаточность
Острая почечная недостаточность Поняття про артеріальний тиск
Поняття про артеріальний тиск Миеломенингоцеле. Знак лимона
Миеломенингоцеле. Знак лимона Үймереттердің ішкі суағары
Үймереттердің ішкі суағары