- Трансплантационный иммунитет

Содержание
- 5. Амебоциты инфильтрировались в аллотрансплантант немертины
- 6. Пересадка имагинальных дисков дрозофилы
- 7. Факторы агрегации губок Amphimedon sp. Clathria sp.
- 8. Слияние (верхний ряд) и отторжение (нижний) колоний у Hydractinia
- 9. Отторжение чужих колоний у гидроида Hydractinia можно объяснить системой всего двух локусов, alr1 и alr2
- 10. Гомофильное связывание изоформ Alr1 и Alr2 обеспечивает сегрегацию клеток Агрегаты клеток СНО, трансфецированных флуоресцентным маркером и
- 11. Полип Stylophora и асцидия Botryllus Морулярные клетки асцидий
- 12. Локус fuhc асцидий содержит 6 генов гистосовместимости?
- 14. Uncle fester fester сfuhc Адгезионные молекулы асцидий
- 15. Трансмиссибельные опухоли
- 16. Лейкемия моллюсков Mya arenaria
- 17. Опухолевые клетки имеют множественные копии транспозона Steamer и демонстрируют общее происхождение
- 18. Гольфстрим разносит опухолевые клетки моллюсков по мировому океану?
- 19. DFTD - Devil Facial Tumour Disease Высокоагрессивная опухоль, возникшая на основе Шванновской клетки, впервые описана в
- 20. Распространение DFTD
- 21. Происхождение DFTD доказывается с помощью иммуногистохимии нерв DFTD Положительное окрашивание на периаксин - белок шванновских клеток
- 22. Клональность DFTD
- 23. Таманские дьяволы действительно высоко изогенны по генам МНС
- 24. Разницы генотипов между больными и здоровыми дьяволами нет
- 25. Отторжение аллографтов у тасманского дьявола
- 26. Венерически передаваемая опухоль собак - CTVT
- 27. CTVT высоко гомозиготна
- 28. Стадия прогрессии CTVT и сниженная экспрессия МНС
- 29. Стадия регрессии CTVT
- 30. Происхождение CTVT и передача от волка собаке
- 31. Саркома Стикера гетерогенна по митохондриальной ДНК
- 33. Многие гены CTVT несут мутации потери функции
- 34. На самом деле, устойчивость DFTD вызвана эпигенети-ческим подавлением экспрессии TAP1, TAP2 и β2-микро-глобулина, что ведет к
- 35. Толерогенное действие клеток DFTD
- 36. Сравнение DFTD и CTVT
- 38. Паразитизм карликовых самцов удильщиков – потенциальная иммунологическая модель?
- 39. Ткани самца и самки удильщиков срастаются
- 41. Скачать презентацию

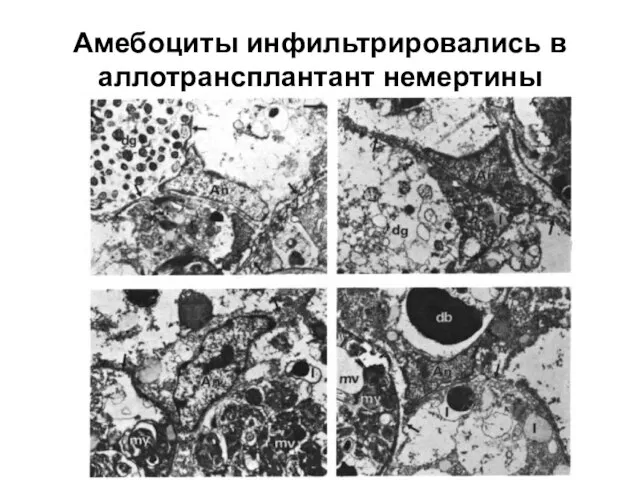
Слайд 5Амебоциты инфильтрировались в аллотрансплантант немертины
Амебоциты инфильтрировались в аллотрансплантант немертины
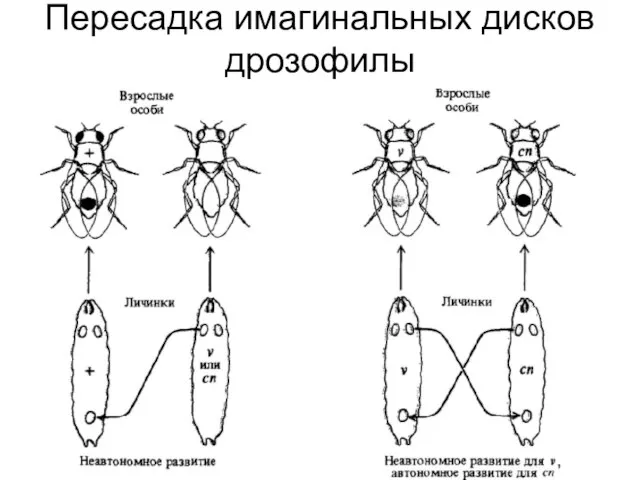
Слайд 6Пересадка имагинальных дисков дрозофилы
Пересадка имагинальных дисков дрозофилы
Слайд 7Факторы агрегации губок
Amphimedon sp.
Clathria sp.
Факторы агрегации губок
Amphimedon sp.
Clathria sp.

Слайд 8Слияние (верхний ряд) и отторжение (нижний) колоний у Hydractinia
Слияние (верхний ряд) и отторжение (нижний) колоний у Hydractinia

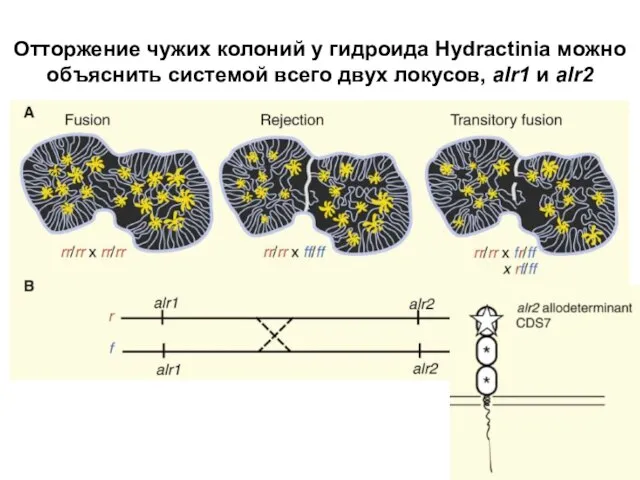
Слайд 9Отторжение чужих колоний у гидроида Hydractinia можно объяснить системой всего двух локусов,
Отторжение чужих колоний у гидроида Hydractinia можно объяснить системой всего двух локусов,
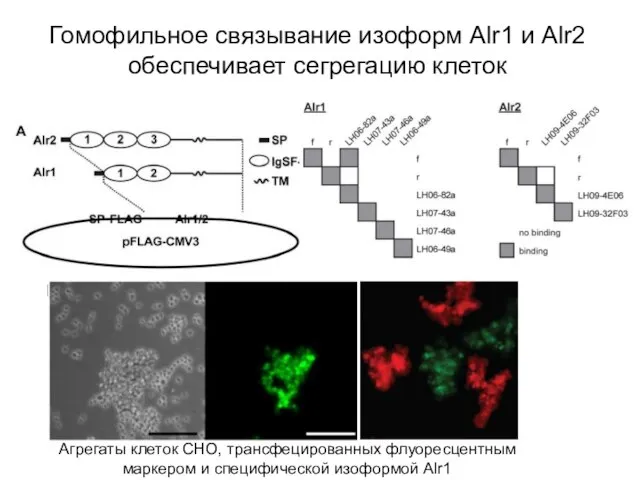
Слайд 10Гомофильное связывание изоформ Alr1 и Alr2 обеспечивает сегрегацию клеток
Агрегаты клеток СНО, трансфецированных
Гомофильное связывание изоформ Alr1 и Alr2 обеспечивает сегрегацию клеток
Агрегаты клеток СНО, трансфецированных
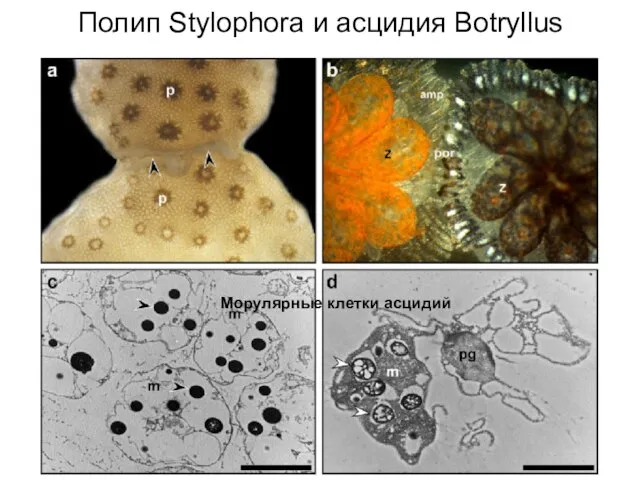
Слайд 11Полип Stylophora и асцидия Botryllus
Морулярные клетки асцидий
Полип Stylophora и асцидия Botryllus
Морулярные клетки асцидий
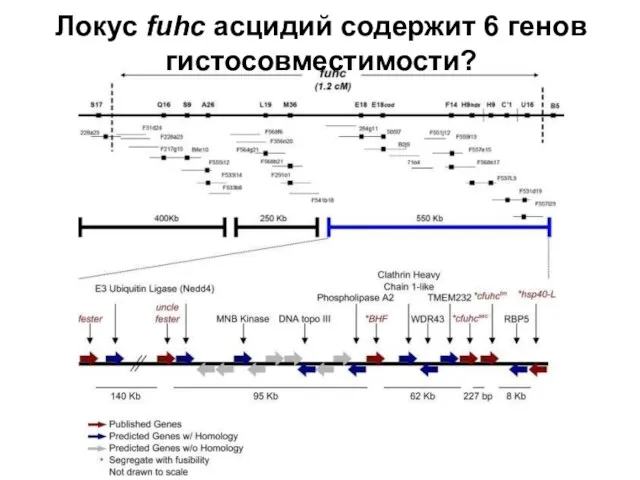
Слайд 12Локус fuhc асцидий содержит 6 генов гистосовместимости?
Локус fuhc асцидий содержит 6 генов гистосовместимости?
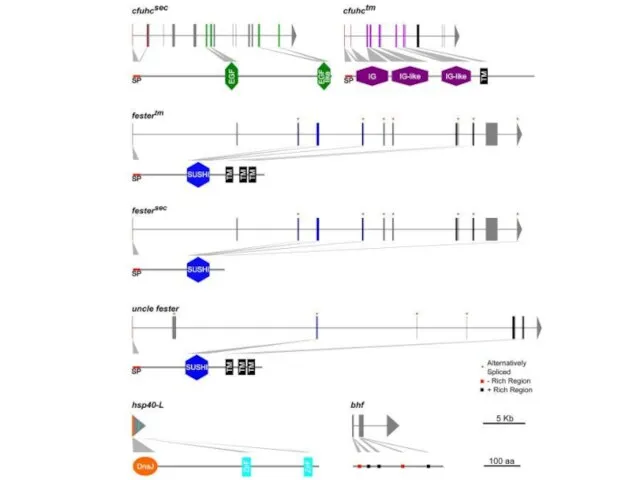
Слайд 14Uncle fester fester сfuhc
Адгезионные молекулы асцидий
Uncle fester fester сfuhc
Адгезионные молекулы асцидий
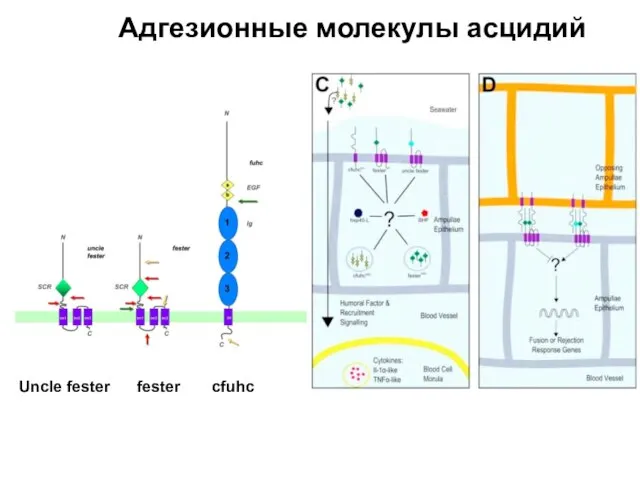
Слайд 15Трансмиссибельные опухоли
Трансмиссибельные опухоли
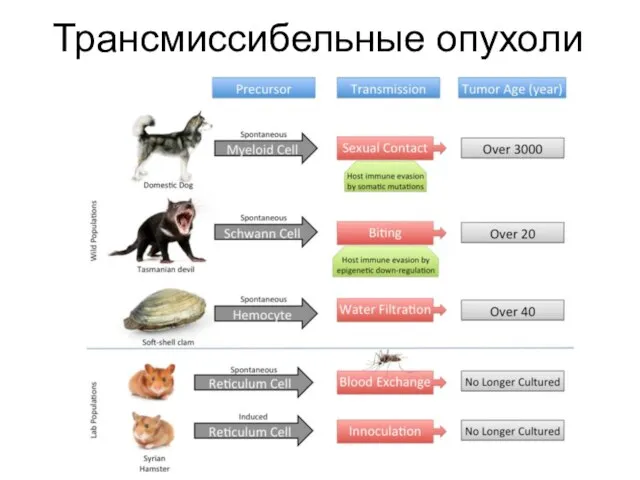
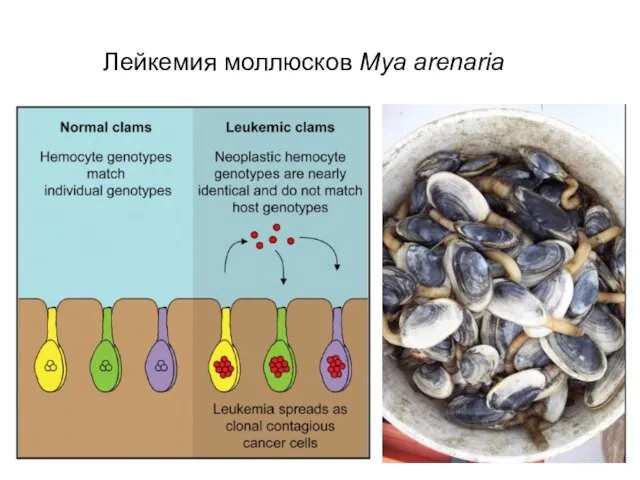
Слайд 16Лейкемия моллюсков Mya arenaria
Лейкемия моллюсков Mya arenaria
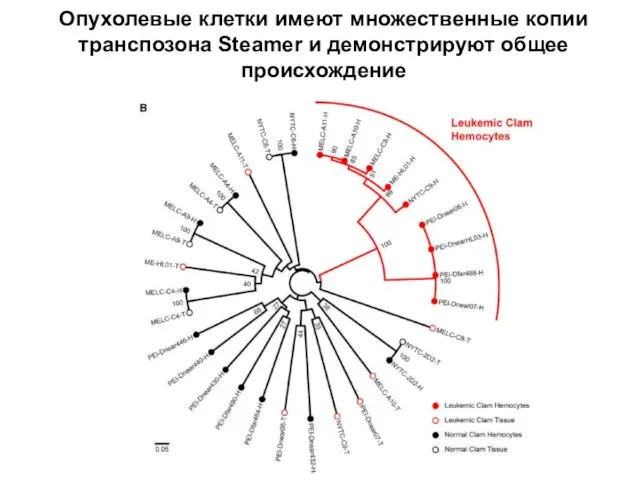
Слайд 17Опухолевые клетки имеют множественные копии транспозона Steamer и демонстрируют общее происхождение
Опухолевые клетки имеют множественные копии транспозона Steamer и демонстрируют общее происхождение
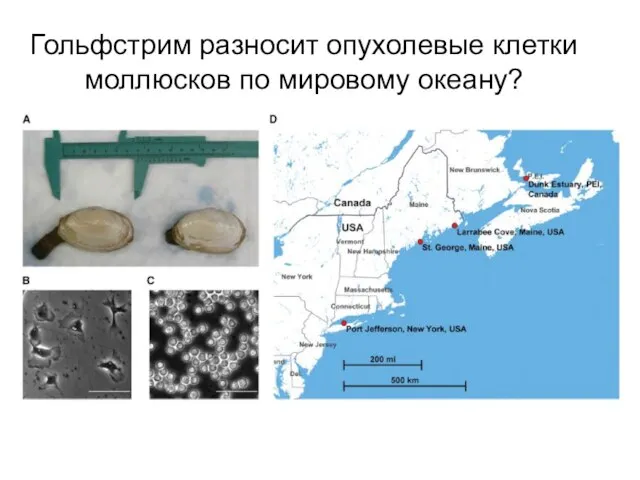
Слайд 18Гольфстрим разносит опухолевые клетки моллюсков по мировому океану?
Гольфстрим разносит опухолевые клетки моллюсков по мировому океану?
Слайд 19DFTD - Devil Facial Tumour Disease
Высокоагрессивная опухоль, возникшая на основе Шванновской клетки,
DFTD - Devil Facial Tumour Disease
Высокоагрессивная опухоль, возникшая на основе Шванновской клетки,
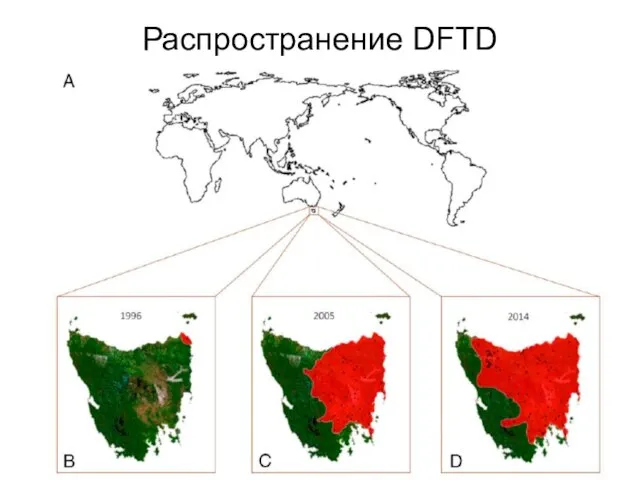
Слайд 20Распространение DFTD
Распространение DFTD
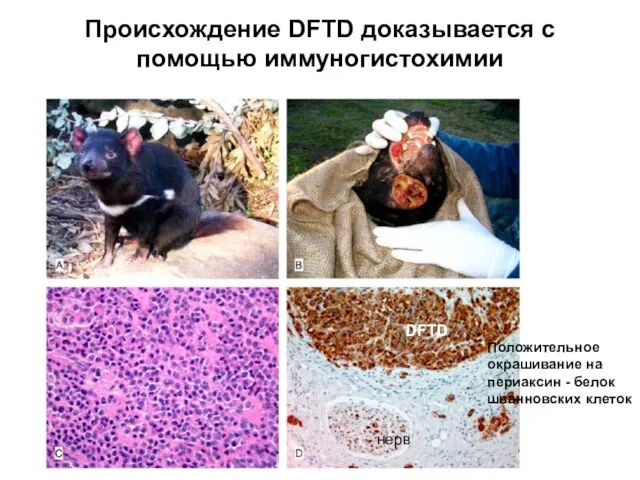
Слайд 21Происхождение DFTD доказывается с помощью иммуногистохимии
нерв
DFTD
Положительное окрашивание на периаксин - белок шванновских
Происхождение DFTD доказывается с помощью иммуногистохимии
нерв
DFTD
Положительное окрашивание на периаксин - белок шванновских
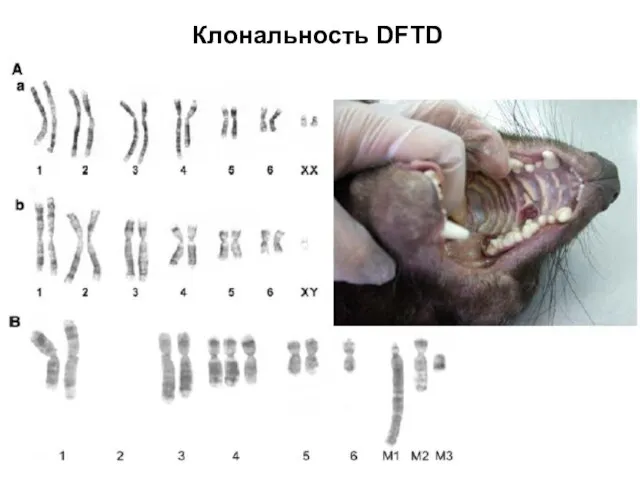
Слайд 22Клональность DFTD
Клональность DFTD
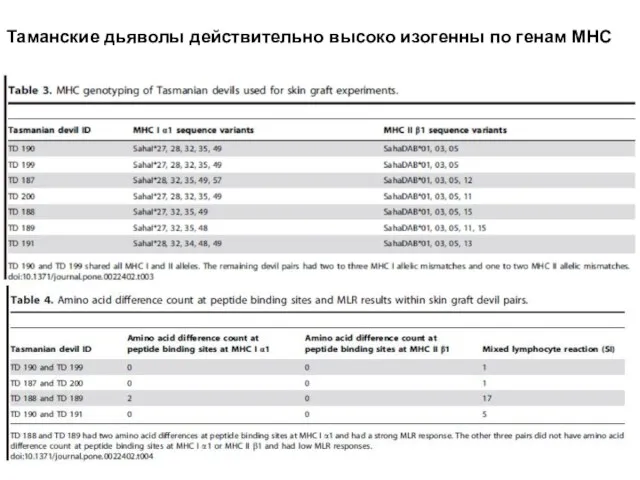
Слайд 23Таманские дьяволы действительно высоко изогенны по генам МНС
Таманские дьяволы действительно высоко изогенны по генам МНС
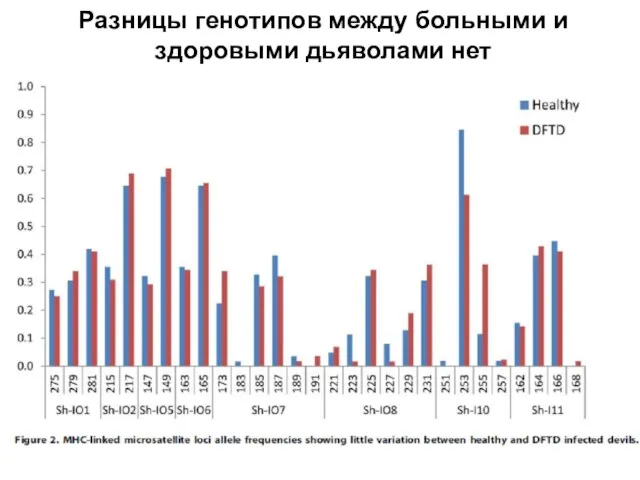
Слайд 24Разницы генотипов между больными и здоровыми дьяволами нет
Разницы генотипов между больными и здоровыми дьяволами нет
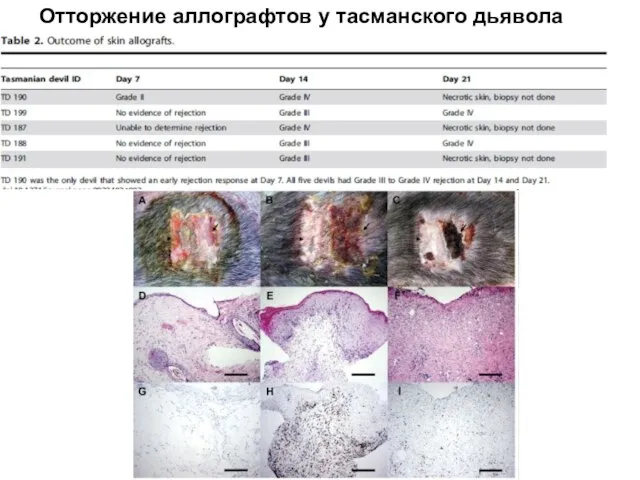
Слайд 25Отторжение аллографтов у тасманского дьявола
Отторжение аллографтов у тасманского дьявола
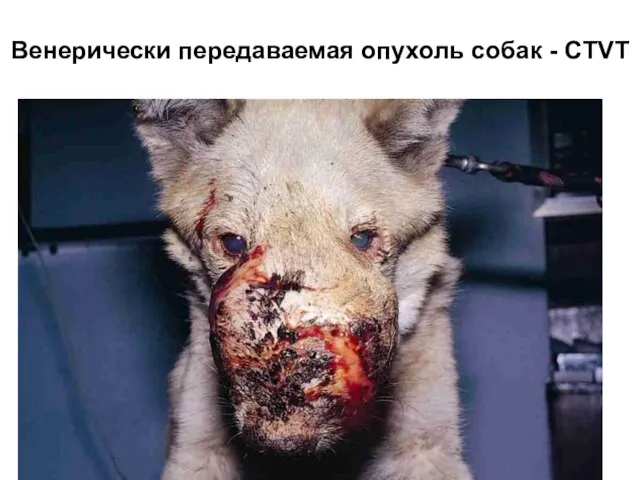
Слайд 26Венерически передаваемая опухоль собак - CTVT
Венерически передаваемая опухоль собак - CTVT
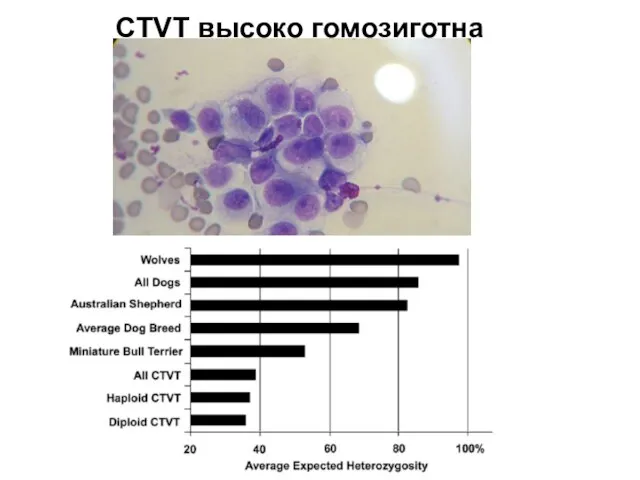
Слайд 27CTVT высоко гомозиготна
CTVT высоко гомозиготна
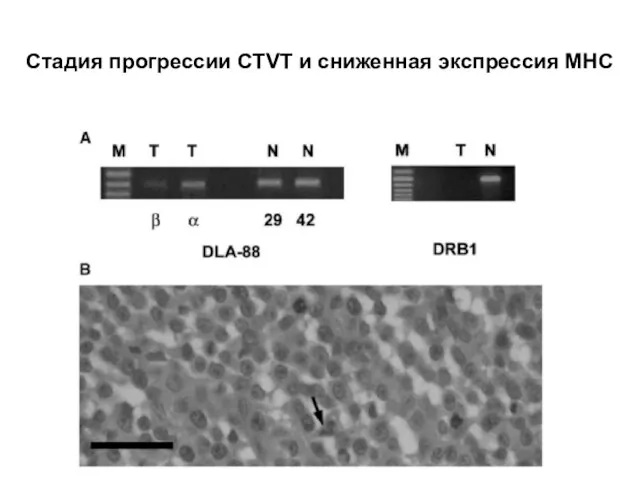
Слайд 28Стадия прогрессии CTVT и сниженная экспрессия МНС
Стадия прогрессии CTVT и сниженная экспрессия МНС
Слайд 29Стадия регрессии CTVT
Стадия регрессии CTVT

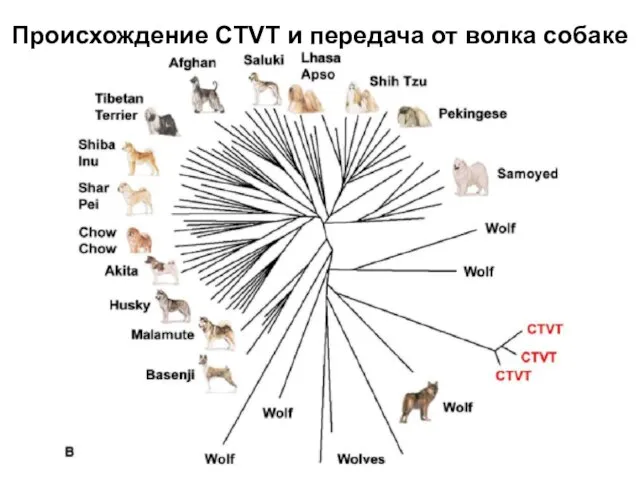
Слайд 30Происхождение CTVT и передача от волка собаке
Происхождение CTVT и передача от волка собаке
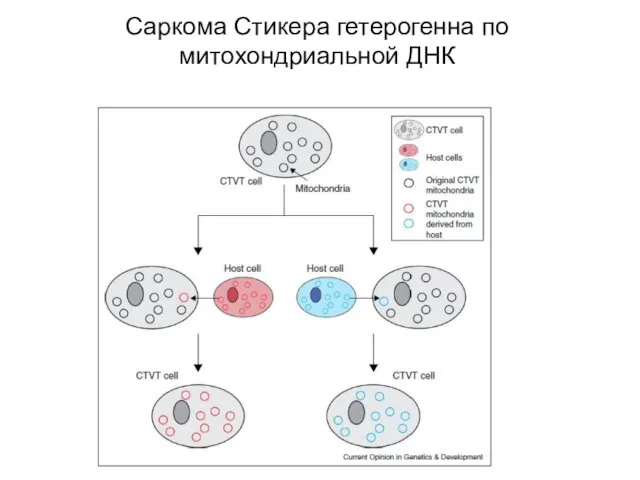
Слайд 31Саркома Стикера гетерогенна по митохондриальной ДНК
Саркома Стикера гетерогенна по митохондриальной ДНК
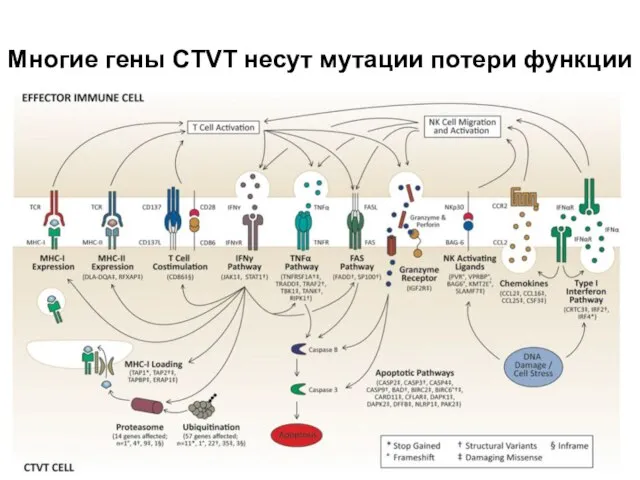
Слайд 33Многие гены CTVT несут мутации потери функции
Многие гены CTVT несут мутации потери функции
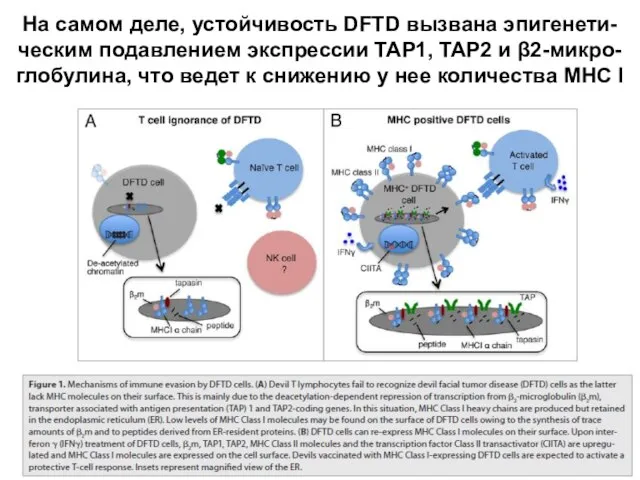
Слайд 34На самом деле, устойчивость DFTD вызвана эпигенети-ческим подавлением экспрессии TAP1, TAP2 и
На самом деле, устойчивость DFTD вызвана эпигенети-ческим подавлением экспрессии TAP1, TAP2 и
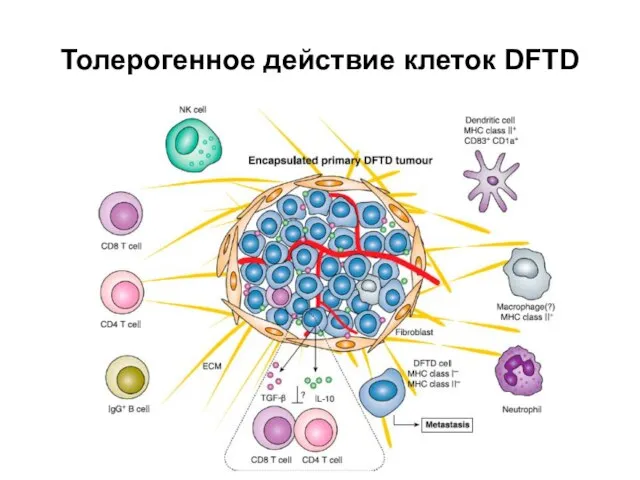
Слайд 35Толерогенное действие клеток DFTD
Толерогенное действие клеток DFTD
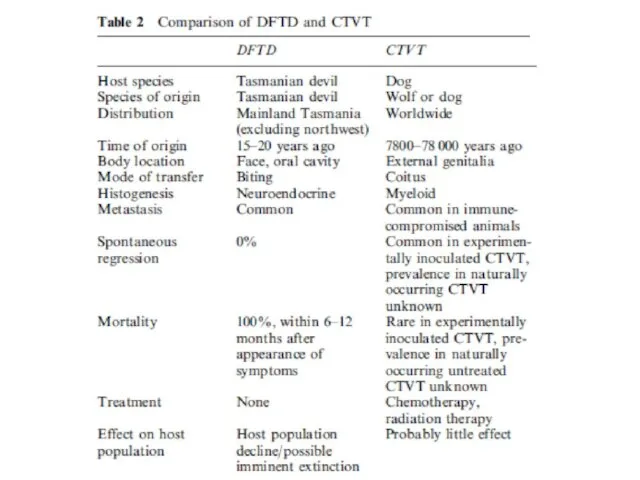
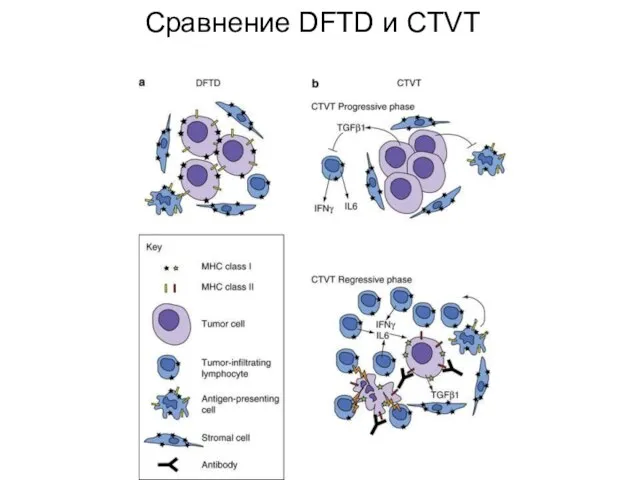
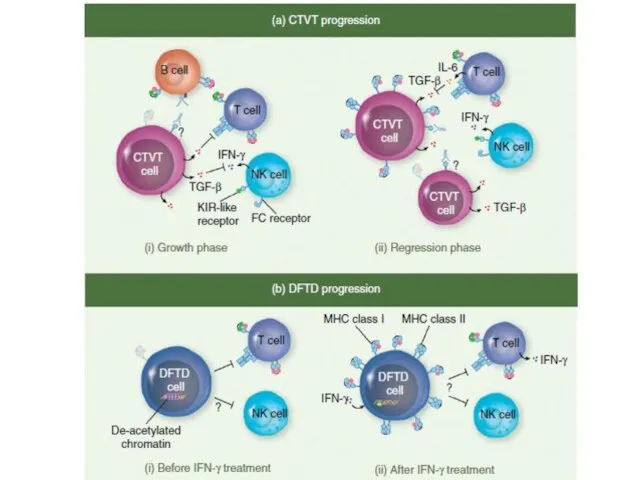
Слайд 36Сравнение DFTD и CTVT
Сравнение DFTD и CTVT
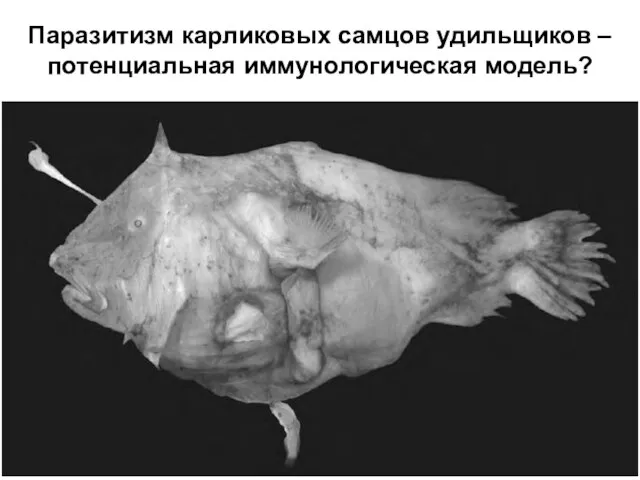
Слайд 38Паразитизм карликовых самцов удильщиков – потенциальная иммунологическая модель?
Паразитизм карликовых самцов удильщиков – потенциальная иммунологическая модель?
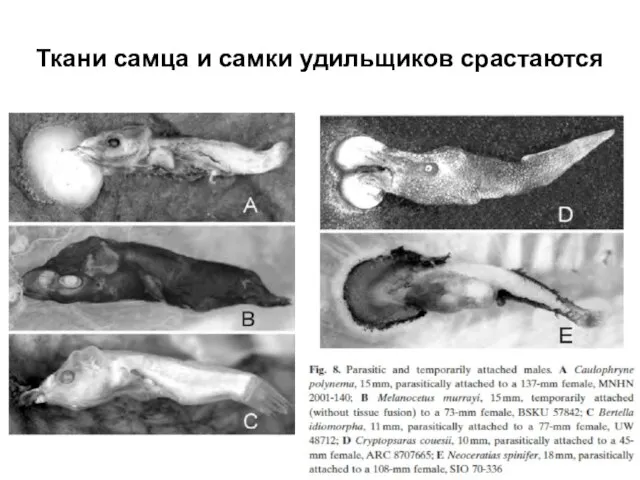
Слайд 39Ткани самца и самки удильщиков срастаются
Ткани самца и самки удильщиков срастаются
 Миотонические синдромы. Миотония Томсена
Миотонические синдромы. Миотония Томсена Психопатологическая семиотика
Психопатологическая семиотика Обморожение. Часть 2
Обморожение. Часть 2 Правила проведения мероприятий по профилактике туберкулеза. Приказ №214
Правила проведения мероприятий по профилактике туберкулеза. Приказ №214 Детские витамины Омега-3
Детские витамины Омега-3 Эргономика труда медицинского персонала: основы правильного перемещения больных после инсульта
Эргономика труда медицинского персонала: основы правильного перемещения больных после инсульта Дисбактериоз у детей
Дисбактериоз у детей Kraevski_1
Kraevski_1 Стандартизация лекарственных растительных средств
Стандартизация лекарственных растительных средств MedPoint v12
MedPoint v12 Воспалительные заболевания органов малого таза. Препараты с противовоспалительным и антиишемическим действием
Воспалительные заболевания органов малого таза. Препараты с противовоспалительным и антиишемическим действием Спецзакаливание как одна из форм оздоровления детей дошкольного возраста из опыта работы инструктора по физическому воспитанию
Спецзакаливание как одна из форм оздоровления детей дошкольного возраста из опыта работы инструктора по физическому воспитанию Холинергические средства (холиномиметики и антихолинэстеразные)
Холинергические средства (холиномиметики и антихолинэстеразные) Депрескрайбинг. Практика снижения дозы и отмены лекарственных средств
Депрескрайбинг. Практика снижения дозы и отмены лекарственных средств Особенности стоматологического статуса населения пожилого возраста
Особенности стоматологического статуса населения пожилого возраста Көз алмасын қозғалтатын бұлшық еттер анатомиясы. IV жұп нервтер
Көз алмасын қозғалтатын бұлшық еттер анатомиясы. IV жұп нервтер Сестринское дело в Республике Казахстан. Технология милосердного ухода, доброты и качества жизни
Сестринское дело в Республике Казахстан. Технология милосердного ухода, доброты и качества жизни История развития хирургии. Основные этапы зарождения и развития хирургии
История развития хирургии. Основные этапы зарождения и развития хирургии Опухоль молочной железы
Опухоль молочной железы Флегмона дна полости рта. Распространенные флегмоны лица и шеи
Флегмона дна полости рта. Распространенные флегмоны лица и шеи Функциональные нарушения и патологии органов речи
Функциональные нарушения и патологии органов речи Ингарон. Лекарственная форма
Ингарон. Лекарственная форма Лучевая диагностика черепно-мозговой травмы. Компьютерная томография
Лучевая диагностика черепно-мозговой травмы. Компьютерная томография Витамины
Витамины Medical academy named after S.I. Georgivsky of vernadsky university
Medical academy named after S.I. Georgivsky of vernadsky university Лекарственные средства, влияющие на функции органов дыхания
Лекарственные средства, влияющие на функции органов дыхания Коклюш
Коклюш Хроническая сердечная недостаточность
Хроническая сердечная недостаточность