- gg

Содержание
- 2. Лимфоциты округлые клетки диаметром 7—9 мкм с круглым или бобовидным ядром и узкой цитоплазмой, бедной цитоплазматическими
- 3. Разные субпопуляции лимфоцитов выполняют различные функции: обеспечение эффективного клеточного иммунитета (в том числе отторжение трансплантата, уничтожение
- 4. По функциональным признакам различают три типа лимфоцитов: B-клеткиB-клетки, T-клеткиB-клетки, T-клетки, NK-клетки. В-лимфоциты распознают чужеродные структуры (антигены
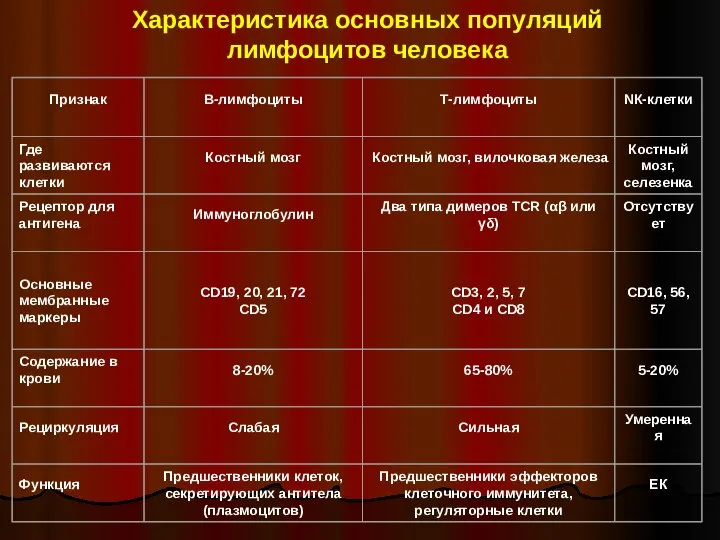
- 5. Характеристика основных популяций лимфоцитов человека
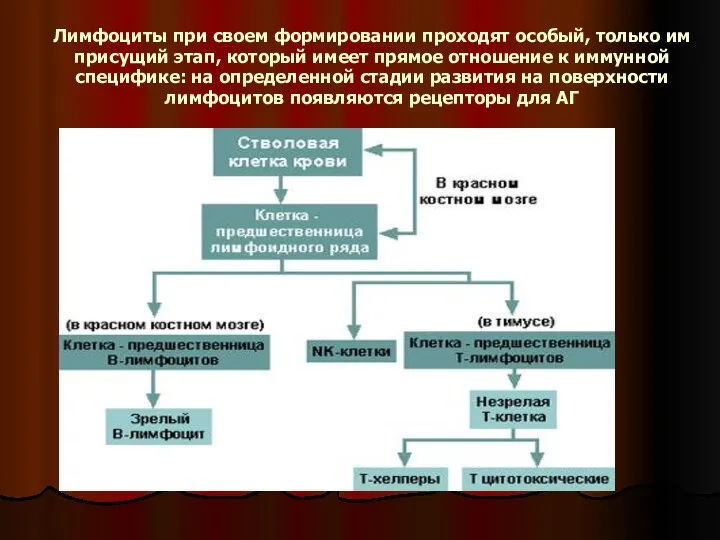
- 7. Лимфоциты при своем формировании проходят особый, только им присущий этап, который имеет прямое отношение к иммунной
- 8. лимфоциты Завершающие развитие в КМ Т-лимфоциты В-лимфоциты Основа рецептора – молекула Ig Основа рецептора -Т-клеточный рецептор,
- 9. Т-лимфоциты
- 10. T-лимфоцит развиваются в тимусе из предшественников — претимоцитов, поступающих из КМ. В тимусе T-л дифференцируются, приобретая
- 11. Существует несколько видов Т-лимфоцитов Т-хелперы Т-киллеры Т-регуляторы Т-клетки памяти
- 12. Т-хелперы(helper — помощник) главной функцией является усиление адаптивного и.о. Активируют Т-киллеры, В-л, моноциты, NK-клетки, презентируя им
- 13. В определении эффекторных механизмов и.о. важный этап- дифференцировка CD4 (T-х)В определении эффекторных механизмов и.о. важный этап-
- 14. Типы Т-хелперов и продуцируемые ими цитокины
- 16. Иммунный ответ развивается в направлении либо TH1-либо TH2-типа. На характер дифференцировки T-х и профиль цитокинов, определяющих
- 17. Цитотоксические Т-л, Т-киллеры (CTL) CD8+лф, осуществляющие лизис, осуществляющие лизис повреждённых клеток собственного организма,поражённых внутриклеточными паразитами (вирусами,
- 18. Механизм действия Т-к непосредственно контактируют с повреждёнными клетками и разрушают их. специфически распознают определённый АГ и
- 19. ХАРАКТЕРИСТИКА ТРЕХ ГЛАВНЫХ ТИПОВ Т-ЛИМФОЦИТОВ
- 20. Клеточный иммунный ответ
- 21. Регуляторные Т-л- CD25+FOXP3+ Т-супрессоры (suppressor T cells, Treg) — центральные регуляторы иммунного ответа. Основная функция —
- 22. Механизмы иммуносупрессии Т-регуляторными клетками Для супрессии и.о. Тreg выделяют цитокины: TGF-beta, IL-10, IFN-gamma, IL-35, а также
- 23. Т-хелперы 17 Th17 - третий тип Т-х, который был открыт в 2005 и назван так из-за
- 24. Т-клетки памяти хранят информацию о ранее действовавших антигенаххранят информацию о ранее действовавших антигенах и формируют вторичный
- 25. В-лимфоциты
- 26. В-лимфоциты (от bursa fabricii птиц, где впервые были обнаружены) — функциональный тип лф, играющих важную роль
- 27. Плазматические клетки- последний этап дифференцировки активированных В-л. несут мало мембранных АТ и способны секретировать растворимые АТ
- 28. Маркеры В-клеток Характерно наличие поверхностных мембрано-связанных АТ, относящихся к классам IgM и IgD. В комплексе с
- 29. Активация В –л-развитие гуморального иммунного ответа
- 30. Роль В-лимфоцитов в презентации антигена В-клетки способны поглощать свои мембранные Ig вместе со связанным ими АГ,
- 31. В-лимфоциты В1(CD5+) В2(CD5-) слизистых оболочек и кожи 2.Секретируют,в основном, IgМ против бактерий 3.На их поверхности экспрессируется
- 32. Естественные киллеры, натуральные киллеры (ЕК,NK) большие гранулярные лимфоциты, обладающие цитотоксичностью против: опухолевых клеток клеток, зараженных вирусами
- 33. фракция клеток, лишенных маркеров Т- и В-лимфоцитов, обозначаемая как 0-клетки способны лизировать определенные клетки-мишени без предварительного
- 34. Рецепторы естественных киллеров Способность NK распознавать «своё» и «чужое» на клетках определяется поверхностными рецепторами. У NK
- 35. Общая схема. Ингибирующий и активирующий сигналы NK. Для предотвращения атаки на неповрежденные клетки на поверхности NK
- 36. Ингибирующий и активирующий сигналы NK
- 37. Механизм действия NK являются цитотоксичными; в их цитоплазме находятся гранулы, содержащие перфорин и протеазы. Перфорин выделяется
- 38. NK перфорин Сериновые эстеразы – гранзимы Хондроэтинсульфат-А белок, обусловливающий образование пор в мембране клеток-мишеней Обусловливающие индукцию
- 39. K Субфракция NK CD16 Рецептор для IgG (FcgRIII) Связывание агрегированных IgG1 и IgG3 Участие в реакциях
- 40. Цитотоксическая функция ЕК-клеток
- 41. Активация лимфоцитов Активация — многозначное понятие. Ее суть сводится к переходу клетки в состояние, с которым
- 42. Апоптоз Апоптоз, или запрограммированная смерть клетки, представляет собой процесс, посредством которого внутренние или внешние факторы, активируя
- 43. Сравнительная характеристика некроза и апоптоза
- 45. Скачать презентацию
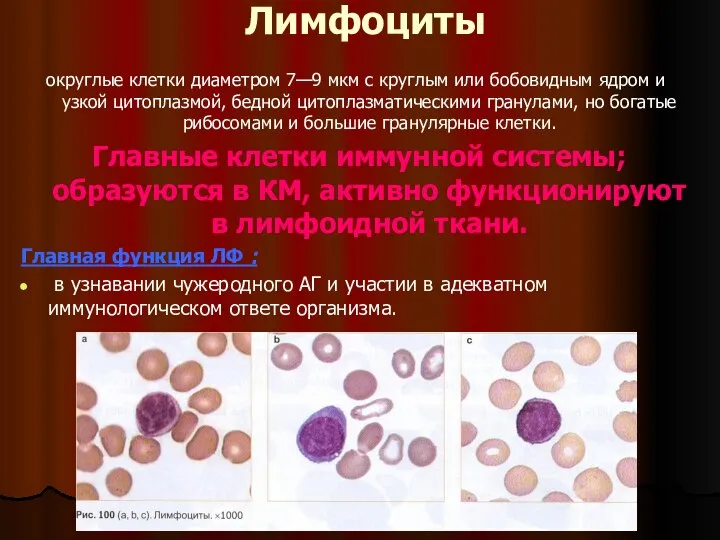
Слайд 3Разные субпопуляции лимфоцитов выполняют различные функции:
обеспечение эффективного клеточного иммунитета (в том числе
Разные субпопуляции лимфоцитов выполняют различные функции:
обеспечение эффективного клеточного иммунитета (в том числе

Слайд 4По функциональным признакам различают три типа лимфоцитов:
B-клеткиB-клетки, T-клеткиB-клетки, T-клетки, NK-клетки.
В-лимфоциты распознают
По функциональным признакам различают три типа лимфоцитов:
B-клеткиB-клетки, T-клеткиB-клетки, T-клетки, NK-клетки.
В-лимфоциты распознают

Слайд 5
Характеристика основных популяций лимфоцитов человека
Характеристика основных популяций лимфоцитов человека
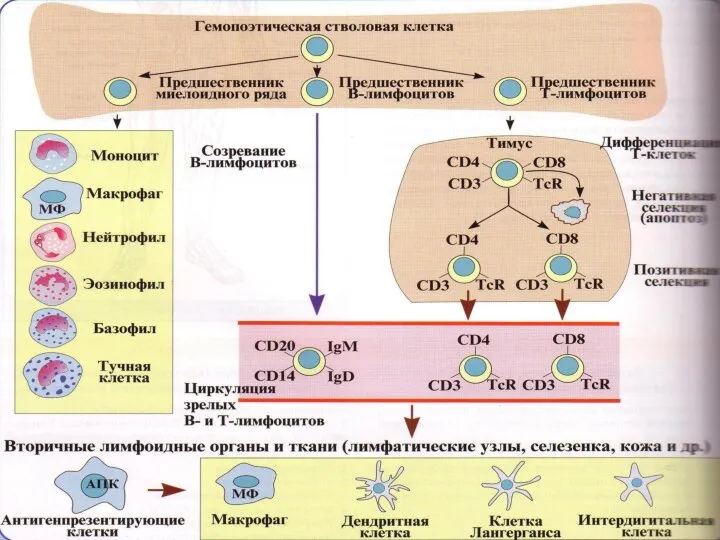
Слайд 7Лимфоциты при своем формировании проходят особый, только им присущий этап, который имеет
Лимфоциты при своем формировании проходят особый, только им присущий этап, который имеет
Слайд 8лимфоциты
Завершающие развитие в КМ
Т-лимфоциты
В-лимфоциты
Основа рецептора – молекула Ig
Основа рецептора -Т-клеточный рецептор,
лимфоциты
Завершающие развитие в КМ
Т-лимфоциты
В-лимфоциты
Основа рецептора – молекула Ig
Основа рецептора -Т-клеточный рецептор,
Слайд 9Т-лимфоциты
Т-лимфоциты
Слайд 10T-лимфоцит
развиваются в тимусе из предшественников — претимоцитов, поступающих из КМ.
В тимусе
T-лимфоцит
развиваются в тимусе из предшественников — претимоцитов, поступающих из КМ.
В тимусе

Слайд 11Существует несколько видов Т-лимфоцитов
Т-хелперы
Т-киллеры
Т-регуляторы
Т-клетки памяти
Существует несколько видов Т-лимфоцитов
Т-хелперы
Т-киллеры
Т-регуляторы
Т-клетки памяти

Слайд 12Т-хелперы(helper — помощник)
главной функцией является усиление адаптивного и.о.
Активируют Т-киллеры, В-л, моноциты,
Т-хелперы(helper — помощник)
главной функцией является усиление адаптивного и.о.
Активируют Т-киллеры, В-л, моноциты,

Слайд 13В определении эффекторных механизмов и.о. важный этап- дифференцировка CD4 (T-х)В определении эффекторных
В определении эффекторных механизмов и.о. важный этап- дифференцировка CD4 (T-х)В определении эффекторных

Слайд 14Типы Т-хелперов и продуцируемые ими цитокины
Типы Т-хелперов и продуцируемые ими цитокины

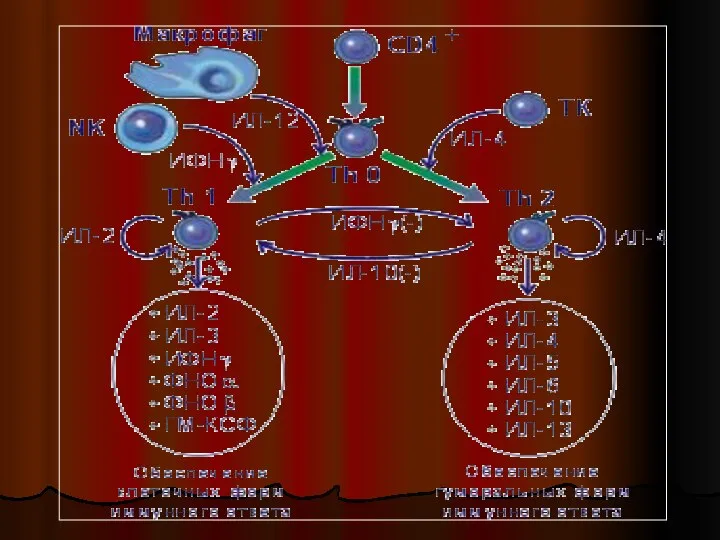
Слайд 16Иммунный ответ развивается в направлении либо TH1-либо TH2-типа.
На характер дифференцировки T-х
Иммунный ответ развивается в направлении либо TH1-либо TH2-типа.
На характер дифференцировки T-х

Слайд 17Цитотоксические Т-л, Т-киллеры (CTL)
CD8+лф, осуществляющие лизис, осуществляющие лизис повреждённых клеток собственного
Цитотоксические Т-л, Т-киллеры (CTL)
CD8+лф, осуществляющие лизис, осуществляющие лизис повреждённых клеток собственного

Слайд 18Механизм действия
Т-к непосредственно контактируют с повреждёнными клетками и разрушают их.
специфически распознают
Механизм действия
Т-к непосредственно контактируют с повреждёнными клетками и разрушают их.
специфически распознают

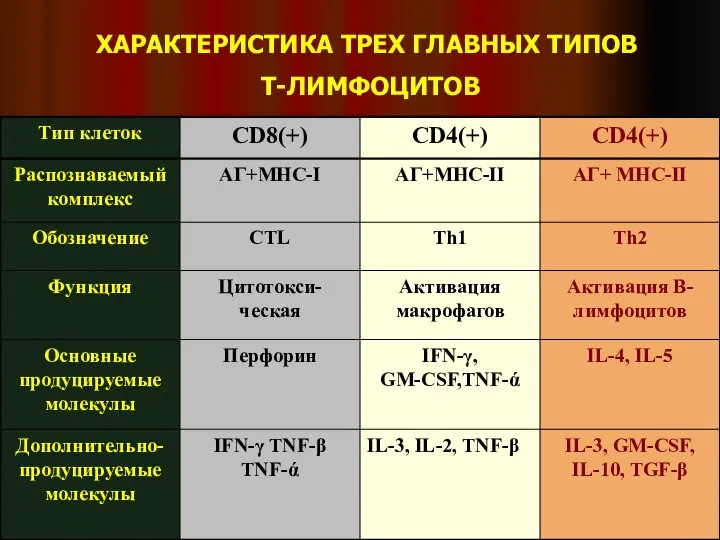
Слайд 19ХАРАКТЕРИСТИКА ТРЕХ ГЛАВНЫХ ТИПОВ
Т-ЛИМФОЦИТОВ
ХАРАКТЕРИСТИКА ТРЕХ ГЛАВНЫХ ТИПОВ
Т-ЛИМФОЦИТОВ
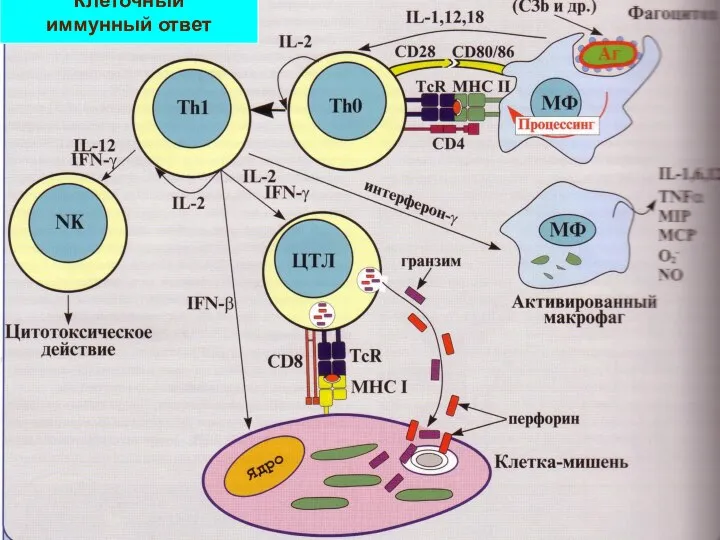
Слайд 20Клеточный
иммунный ответ
Клеточный
иммунный ответ
Слайд 21Регуляторные Т-л- CD25+FOXP3+
Т-супрессоры (suppressor T cells, Treg) — центральные регуляторы иммунного
Регуляторные Т-л- CD25+FOXP3+
Т-супрессоры (suppressor T cells, Treg) — центральные регуляторы иммунного

Слайд 22Механизмы иммуносупрессии Т-регуляторными клетками
Для супрессии и.о. Тreg выделяют цитокины: TGF-beta, IL-10, IFN-gamma,
Механизмы иммуносупрессии Т-регуляторными клетками
Для супрессии и.о. Тreg выделяют цитокины: TGF-beta, IL-10, IFN-gamma,

Слайд 23Т-хелперы 17
Th17 - третий тип Т-х, который был открыт в 2005 и
Т-хелперы 17
Th17 - третий тип Т-х, который был открыт в 2005 и

Слайд 24Т-клетки памяти
хранят информацию о ранее действовавших антигенаххранят информацию о ранее действовавших антигенах
Т-клетки памяти
хранят информацию о ранее действовавших антигенаххранят информацию о ранее действовавших антигенах

Слайд 25В-лимфоциты
В-лимфоциты
Слайд 26
В-лимфоциты
(от bursa fabricii птиц, где впервые были обнаружены) — функциональный тип
В-лимфоциты
(от bursa fabricii птиц, где впервые были обнаружены) — функциональный тип

Слайд 27
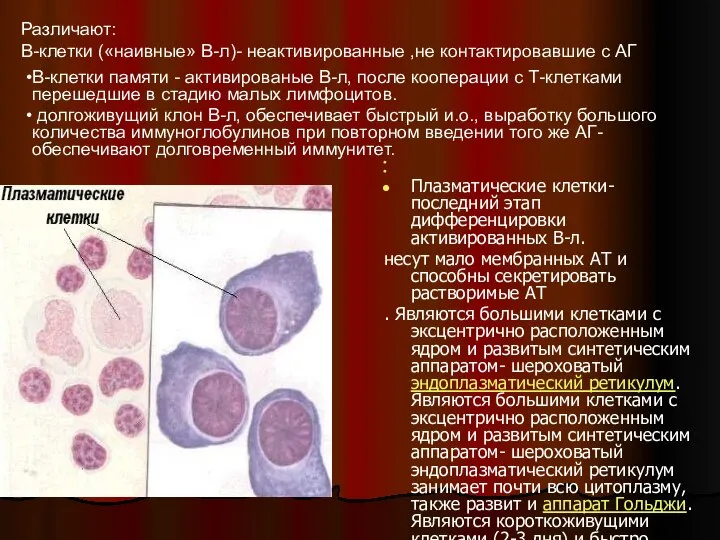
Плазматические клетки- последний этап дифференцировки активированных В-л.
несут мало мембранных АТ и
Плазматические клетки- последний этап дифференцировки активированных В-л.
несут мало мембранных АТ и
Слайд 28Маркеры В-клеток
Характерно наличие поверхностных мембрано-связанных АТ, относящихся к классам IgM и IgD.
В
Маркеры В-клеток
Характерно наличие поверхностных мембрано-связанных АТ, относящихся к классам IgM и IgD.
В

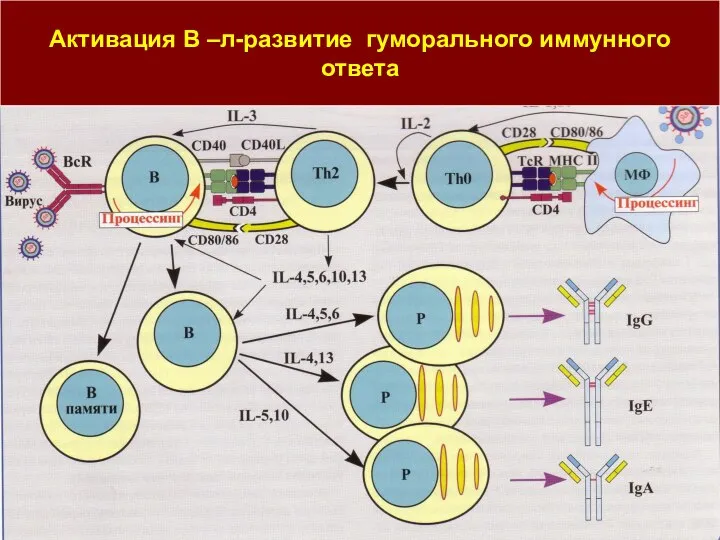
Слайд 29Активация В –л-развитие гуморального иммунного ответа
Активация В –л-развитие гуморального иммунного ответа
Слайд 30Роль В-лимфоцитов в презентации антигена
В-клетки способны поглощать свои мембранные Ig вместе со
Роль В-лимфоцитов в презентации антигена
В-клетки способны поглощать свои мембранные Ig вместе со

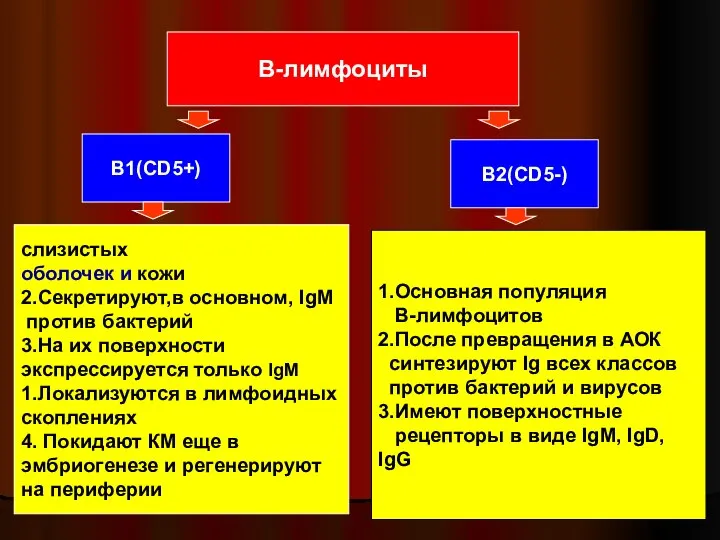
Слайд 31В-лимфоциты
В1(CD5+)
В2(CD5-)
слизистых
оболочек и кожи
2.Секретируют,в основном, IgМ
против бактерий
3.На их поверхности
экспрессируется только
В-лимфоциты
В1(CD5+)
В2(CD5-)
слизистых
оболочек и кожи
2.Секретируют,в основном, IgМ
против бактерий
3.На их поверхности
экспрессируется только
Слайд 32Естественные киллеры, натуральные киллеры (ЕК,NK)
большие гранулярные лимфоциты, обладающие цитотоксичностью против:
опухолевых клеток
Естественные киллеры, натуральные киллеры (ЕК,NK)
большие гранулярные лимфоциты, обладающие цитотоксичностью против:
опухолевых клеток

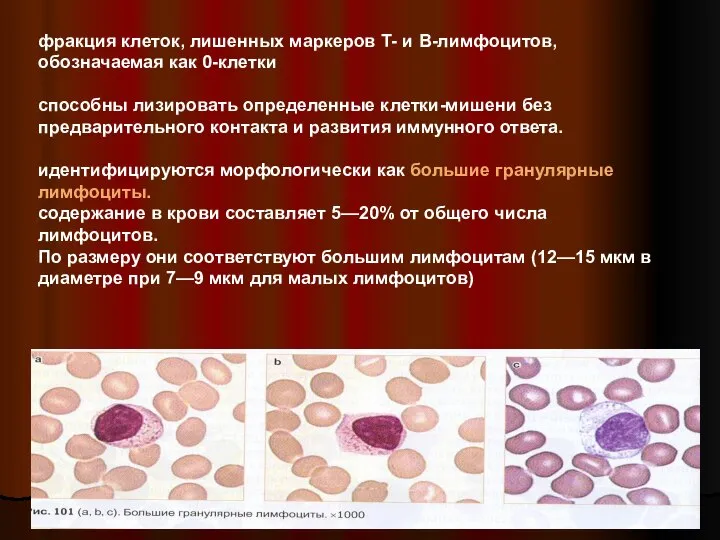
Слайд 33фракция клеток, лишенных маркеров Т- и В-лимфоцитов, обозначаемая как 0-клетки
способны лизировать
фракция клеток, лишенных маркеров Т- и В-лимфоцитов, обозначаемая как 0-клетки
способны лизировать
Слайд 34Рецепторы естественных киллеров
Способность NK распознавать «своё» и «чужое» на клетках определяется поверхностными
Рецепторы естественных киллеров
Способность NK распознавать «своё» и «чужое» на клетках определяется поверхностными

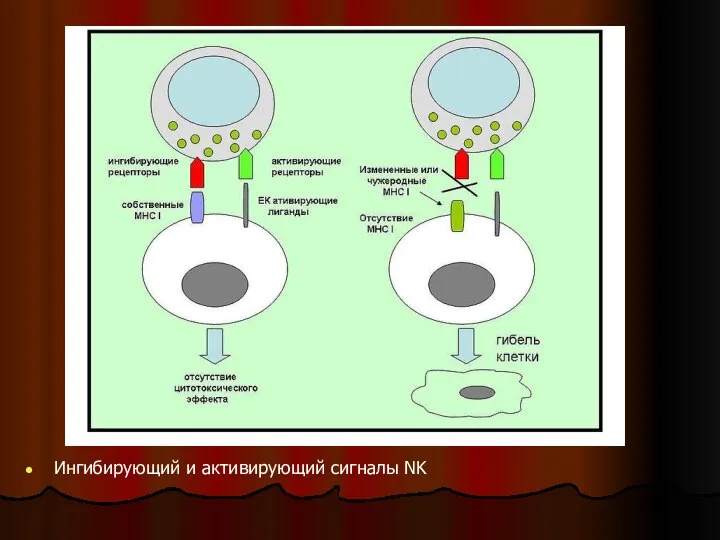
Слайд 35Общая схема. Ингибирующий и активирующий сигналы NK.
Для предотвращения атаки на неповрежденные клетки
Общая схема. Ингибирующий и активирующий сигналы NK.
Для предотвращения атаки на неповрежденные клетки

Слайд 36Ингибирующий и активирующий сигналы NK
Ингибирующий и активирующий сигналы NK
Слайд 37Механизм действия
NK являются цитотоксичными;
в их цитоплазме находятся гранулы, содержащие перфорин и
Механизм действия
NK являются цитотоксичными;
в их цитоплазме находятся гранулы, содержащие перфорин и

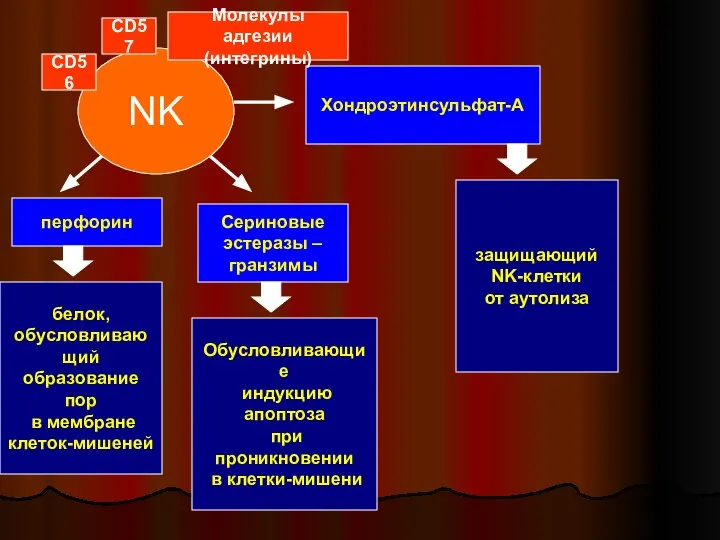
Слайд 38NK
перфорин
Сериновые
эстеразы –
гранзимы
Хондроэтинсульфат-А
белок,
обусловливающий
образование пор
в мембране
клеток-мишеней
Обусловливающие
индукцию
NK
перфорин
Сериновые
эстеразы –
гранзимы
Хондроэтинсульфат-А
белок,
обусловливающий
образование пор
в мембране
клеток-мишеней
Обусловливающие
индукцию
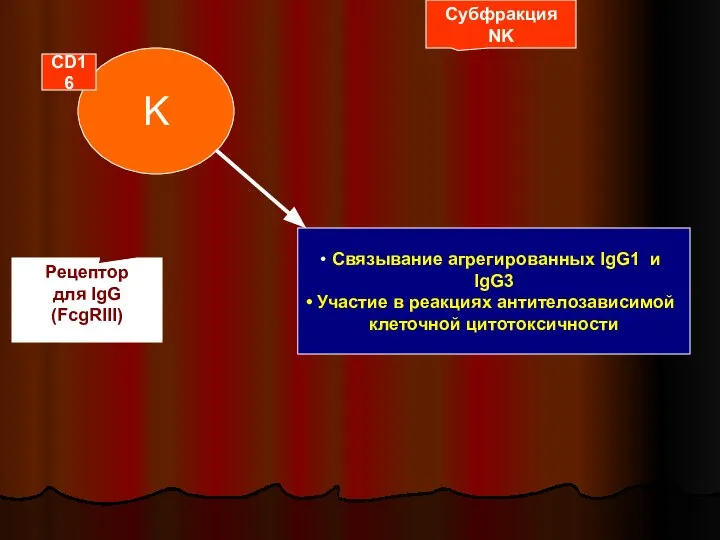
Слайд 39K
Субфракция
NK
CD16
Рецептор
для IgG
(FcgRIII)
Связывание агрегированных IgG1 и IgG3
Участие в реакциях антителозависимой
клеточной цитотоксичности
K
Субфракция
NK
CD16
Рецептор
для IgG
(FcgRIII)
Связывание агрегированных IgG1 и IgG3
Участие в реакциях антителозависимой
клеточной цитотоксичности
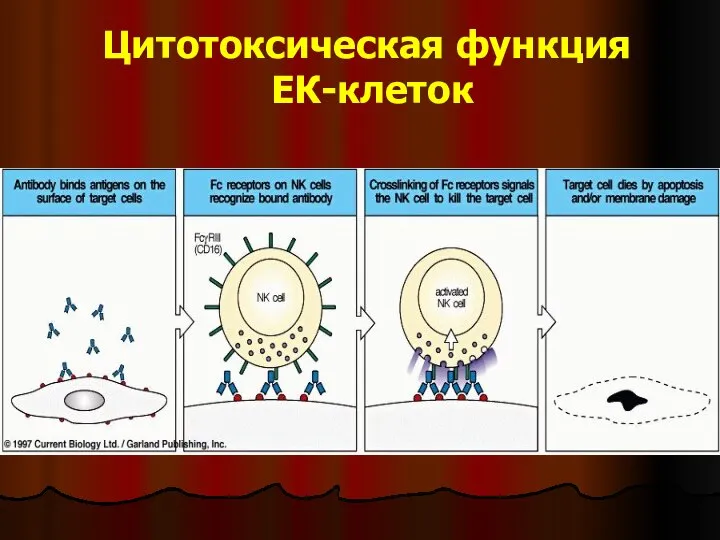
Слайд 40Цитотоксическая функция
ЕК-клеток
Цитотоксическая функция
ЕК-клеток
Слайд 41Активация лимфоцитов
Активация — многозначное понятие. Ее суть сводится к переходу клетки
Активация лимфоцитов
Активация — многозначное понятие. Ее суть сводится к переходу клетки

Слайд 42Апоптоз
Апоптоз, или запрограммированная смерть клетки, представляет собой процесс, посредством которого внутренние или
Апоптоз
Апоптоз, или запрограммированная смерть клетки, представляет собой процесс, посредством которого внутренние или

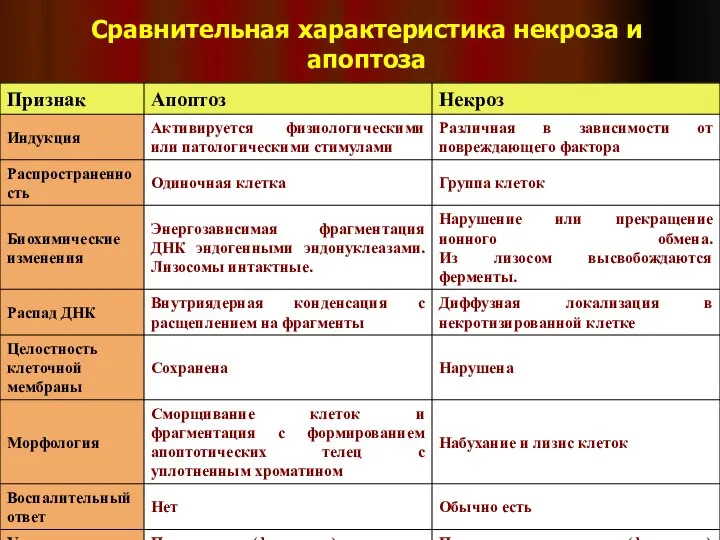
Слайд 43Сравнительная характеристика некроза и апоптоза
Сравнительная характеристика некроза и апоптоза
 Кровь и лимфа. Гемопоэз
Кровь и лимфа. Гемопоэз Профилактика первичная и вторичная в амбулаторно-поликлиническом звене здравоохранения. Диспансерный метод терапевта
Профилактика первичная и вторичная в амбулаторно-поликлиническом звене здравоохранения. Диспансерный метод терапевта Учреждение Частная многопрофильная клиника Венера
Учреждение Частная многопрофильная клиника Венера Вред курения
Вред курения Сильнодействующие ядовитые вещества и последствия их применения
Сильнодействующие ядовитые вещества и последствия их применения Методы обследования при заболеваниях органов дыхания
Методы обследования при заболеваниях органов дыхания срс — копия
срс — копия Грибы детям. Плюсы и минусы
Грибы детям. Плюсы и минусы 病例展示
病例展示 Гигиена. Личная гигиена
Гигиена. Личная гигиена Морфология соединений костей. Биомеханика суставов. Изменения соединений костей
Морфология соединений костей. Биомеханика суставов. Изменения соединений костей Микард-Лана технология телеметрии ЭКГ 3.0. Функциональная диагностика в поликлиниках и больницах
Микард-Лана технология телеметрии ЭКГ 3.0. Функциональная диагностика в поликлиниках и больницах Пищевые отравления
Пищевые отравления Всё ли мы знаем о ГЭРБ
Всё ли мы знаем о ГЭРБ Чистые руки. Правила мытья рук
Чистые руки. Правила мытья рук Сестринский уход при рожистом воспалении
Сестринский уход при рожистом воспалении Лечение повреждений вен. Методы восстановления
Лечение повреждений вен. Методы восстановления Ателектазы у детей
Ателектазы у детей ЧУЗ Медико-санитарная часть. Клиника высокотехнологичной медицины на службе вашего здоровья
ЧУЗ Медико-санитарная часть. Клиника высокотехнологичной медицины на службе вашего здоровья Анатомо-физиологические особенности детей дошкольного возраста
Анатомо-физиологические особенности детей дошкольного возраста Игровой массаж и самомассаж в работе с детьми ОВЗ
Игровой массаж и самомассаж в работе с детьми ОВЗ Физиология и патология менопаузального периода
Физиология и патология менопаузального периода Принципы оказания психиатрической помощи
Принципы оказания психиатрической помощи Аутизм, РАС – биокоррекция. Занятие 2
Аутизм, РАС – биокоррекция. Занятие 2 Картина здоровья
Картина здоровья Периоды детского возраста
Периоды детского возраста The correlation coe cient between DOCK1 and claudin-1
The correlation coe cient between DOCK1 and claudin-1 Рентгенодиагностика неопухолевых заболеваний пищевода
Рентгенодиагностика неопухолевых заболеваний пищевода