Организм как континуум нормальных и иммунных функций, взаимно замещающих друг друга (иммунология, лекция 3)
- Организм как континуум нормальных и иммунных функций, взаимно замещающих друг друга (иммунология, лекция 3)

Содержание
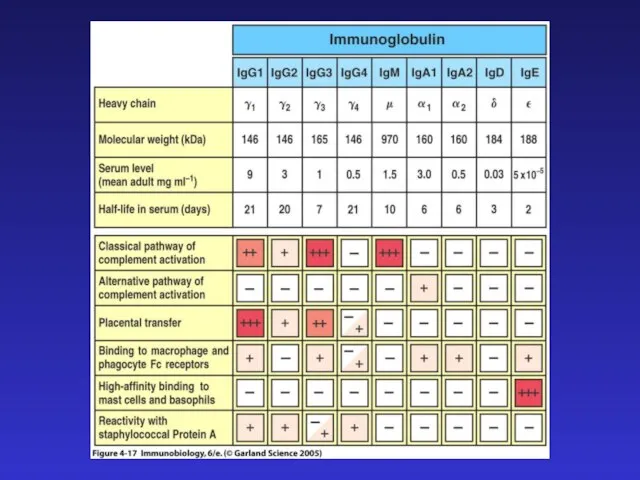
- 10. J-цепи в IgM и IgA позволяют этим иммуноглобулинам существовать в секретируемой форме пентамера и димера соответственно.
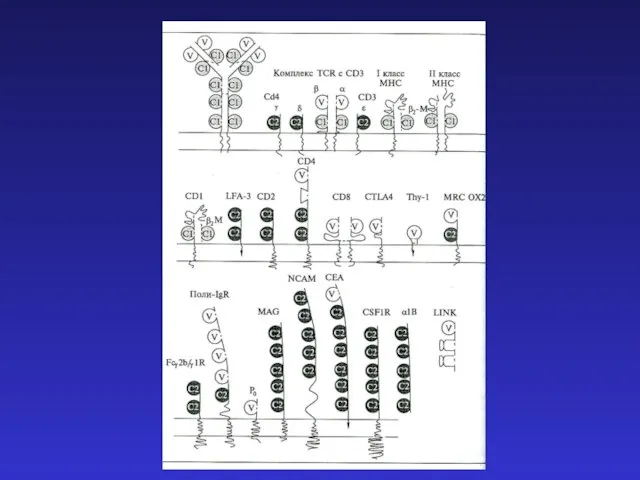
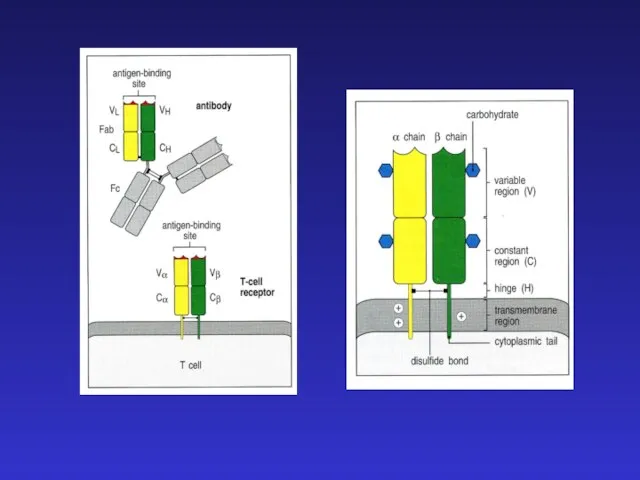
- 24. Существует сходство между процессами реаранжировки цепей Ig и TCR: а) легких цепей иммуноглобулинов и α-цепи TCR
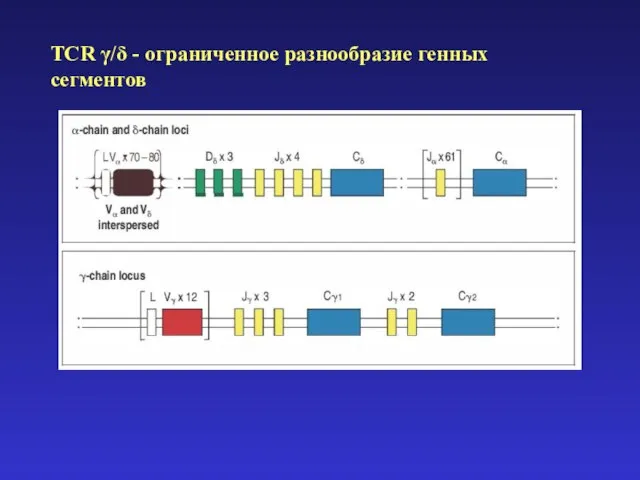
- 25. Четыре главных процесса, формирующих разнообразие антигенспецифических рецепторов: 1) комбинаторное разнообразие в результате рекомбинации V-(D)-J 2) добавление
- 27. Скачать презентацию






Слайд 10J-цепи в IgM и IgA позволяют этим иммуноглобулинам
существовать в секретируемой форме пентамера
J-цепи в IgM и IgA позволяют этим иммуноглобулинам
существовать в секретируемой форме пентамера












Слайд 24Существует сходство между процессами реаранжировки
цепей Ig и TCR:
а) легких цепей иммуноглобулинов и
Существует сходство между процессами реаранжировки
цепей Ig и TCR:
а) легких цепей иммуноглобулинов и

Слайд 25Четыре главных процесса, формирующих разнообразие
антигенспецифических рецепторов:
1) комбинаторное разнообразие в результате рекомбинации
Четыре главных процесса, формирующих разнообразие
антигенспецифических рецепторов:
1) комбинаторное разнообразие в результате рекомбинации

 reabilitatsia_zabolevaniy_organov_dykhania (1)
reabilitatsia_zabolevaniy_organov_dykhania (1) Ультразвуковая допплерография в стоматологии
Ультразвуковая допплерография в стоматологии Депрессия (от латинского слова depressio - подавление)
Депрессия (от латинского слова depressio - подавление) Нарушение осанки и плоскостопие
Нарушение осанки и плоскостопие Курение как социальное зло в наше время
Курение как социальное зло в наше время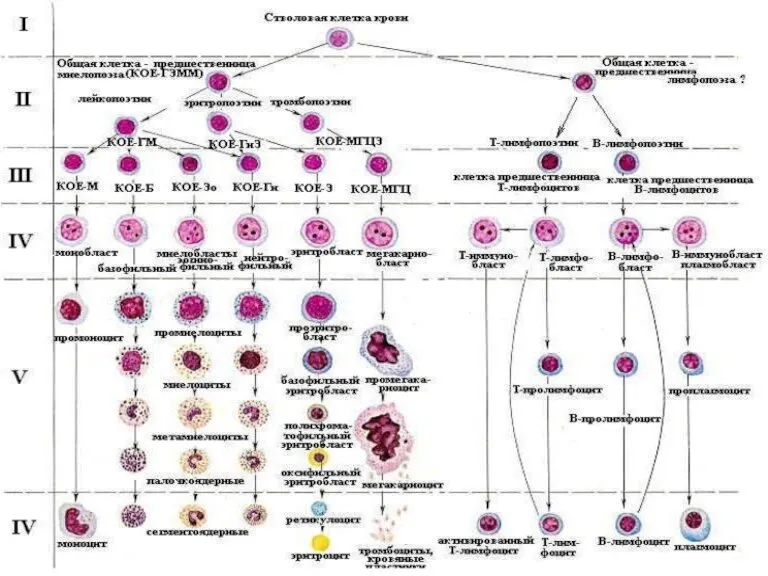 Болезни системы крови
Болезни системы крови Heart failure slideset
Heart failure slideset Хирургическая онкология
Хирургическая онкология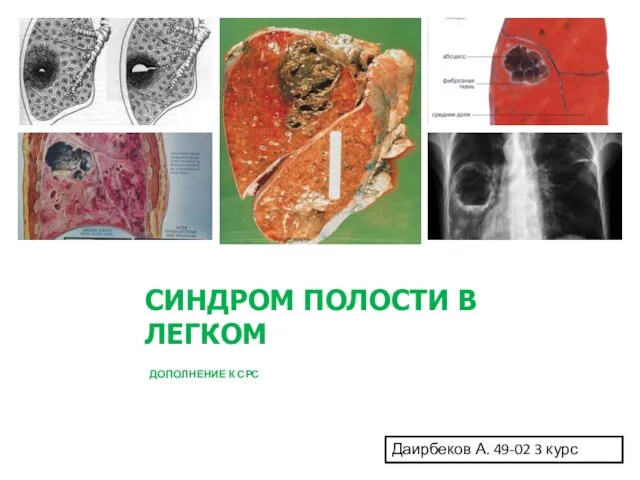 Синдром полости в легком
Синдром полости в легком Тема Роды
Тема Роды ВИЧ и его профилактика
ВИЧ и его профилактика Принципы современной защиты от гриппа и ОРЗ и аптечные продажи
Принципы современной защиты от гриппа и ОРЗ и аптечные продажи СПИД
СПИД Цереброваскулярлы аурулар
Цереброваскулярлы аурулар Первая помощь при кровотечениях
Первая помощь при кровотечениях Чесотка. Чесоточный клещ
Чесотка. Чесоточный клещ Заболевания нервной системы и их профилактика
Заболевания нервной системы и их профилактика Клеточные элементы ликвора
Клеточные элементы ликвора Дисбиотические патологические процессы
Дисбиотические патологические процессы ВИЧ-инфекция
ВИЧ-инфекция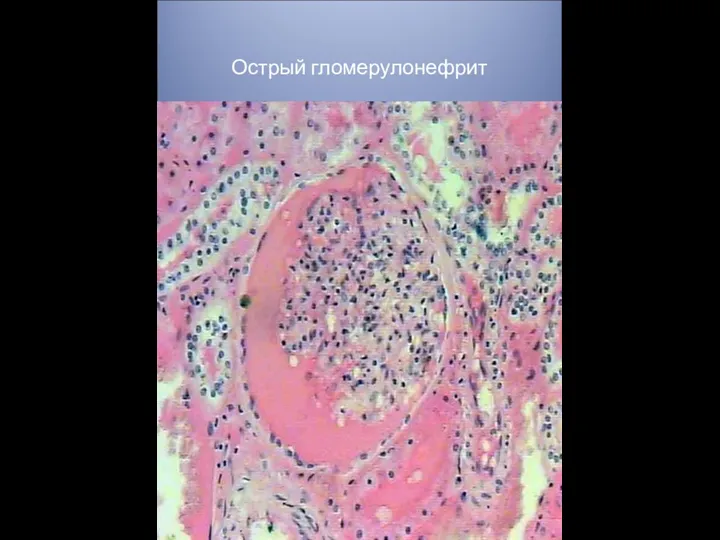 Острый гломерулонефрит
Острый гломерулонефрит Медицинские учреждения
Медицинские учреждения Травматический шок
Травматический шок Сознание
Сознание Природные антибактериальные средства
Природные антибактериальные средства Диагностика хронической недостаточности сердца
Диагностика хронической недостаточности сердца Особенности диагностики. Оказание неотложной помощи. Интенсивная терапия
Особенности диагностики. Оказание неотложной помощи. Интенсивная терапия Аналар мен балалар денсаулығын қорғау
Аналар мен балалар денсаулығын қорғау